Effects of treadmill exercise on the anxiety-like behavior through modulation of GSK3β/β-catenin signaling in the maternal separation rat pup
Article information

Abstract
Maternal separation causes depression and anxiety. Exercise ameliorates maternal separation-induced depression. In this study, we investigated the effect of treadmill exercise on anxiety-like behavior in relation with glycogen synthase kinase 3 beta (GSK3β)/β-catenin pathway using maternal separation rat pups. For this study, elevated plus maze test, immunohistochemistry for serotonin (5-hydroxytryptamine, 5-HT), tryptophan hydroxylase (TPH), and western blot for total GSK3β (t-GSK3β), phosphorylated GSK3β (p-GSK3β), total β-catenin (t-β-catenin), and phosphorylated β-catenin (p-β-catenin) were conducted. The rat pups in the exercise groups were scheduled to run on a treadmill for 30 min once a day for 10 days, starting on postnatal day 21. For the rat pups in the fluoxetine-treated group, fluoxetine was orally administrated once a day for 10 consecutive days, starting on postnatal day 21. Anxiety-like behavior was appeared in the rat pups by maternal separation. Maternal separation suppressed 5-HT and TPH expression in the dorsal raphe. Maternal separation suppressed phosphorylation of GSK3β and increased phosphorylation of β-catenin in the hippocampus. However, treadmill exercise and fluoxetine treatment alleviated anxiety and increased 5-HT and TPH expression in the dorsal raphe. Treadmill exercise and fluoxetine treatment also enhanced GSK3β phosphorylation and suppressed β-catenin phosphorylation in the hippocampus. In this study, alleviating effect of treadmill exercise on maternal separation-induced anxiety appeared through enhancing 5-HT expression and GSK3β phosphorylation, and then inhibiting β-catenin phosphorylation. These results showed that treadmill exercise relieves anxiety through GSK3β/β-catenin pathway. Treadmill exercise showed similar ameliorating effect on anxiety-like behavior as fluoxetine.
INTRODUCTION
Close relationship with parents is important factor for the development of offspring. Maternal separation causes emotional, behavioral, cognitive, and socialization problems to offspring. Maternal separation is one of the animal models of depression, and depression induces a functional alteration of various neurotransmitters (Baek et al., 2012; Yadid et al., 2000).
Among the neurotransmitters related with depression and anxiety, serotonin (5-hydroxy trigramamine, 5-HT) is involved in feeding, sleep, diet, sleep, and mood control (Bannai et al., 2007; Ji et al., 2017). Tryptophan hydroxylase (TPH) is known as a speed-limiting enzyme for the production of 5-HT, and the level of TPH has been used as an indicator of 5-HT synthesis. Depression is associated with down-regulation of 5-HT in the dorsal raphe, and 5-HT controls neural stabilization and alleviates depressive behavior (Baek et al., 2012; Roh et al., 2016).
5-HT regulates glycogen synthase kinase 3 beta (GSK3β), and GSK3β is serine/threonine kinase which modulates glycogen biosynthesis (Latapy et al., 2012). Impaired GSK3β activity occurs under the conditions of dysregulation in serotonergic activity, such as mood disorders (Li et al., 2004). Inhibition of GSK3β exerted an anti-depressive effect, and then GSK3β inhibitors have been considered as the antidepressants (Kaidanovich-Beilin et al., 2004). GSK3β is associated with the serotonin-sensitive anxiety and social behavior (Latapy et al., 2012).
The transcription factor β-catenin is the marker of GSK3β inactivation, because cytoplasmic level of β-catenin is increased by inhibition of the GSK3β (Gould et al., 2004). GSK3β/β-catenin signaling plays an important role in stress-related depression, and this pathway contributes to the anti-depressive effect in the mood regulation brain region (Chen et al., 2012).
Exercise is known to increased brain 5-HT level and exercise acts as noninvasive treatment for depression (Baek et al., 2012; Ji et al., 2017; Lee and Baek, 2017). Fluoxetine is the most widely used antidepressant acting as one of the serotonin reuptake inhibitors (SSRIs) (Ferguson, 2001). In the present study, we investigated the effect of treadmill exercise on the maternal separation-induced anxiety in relation with GSK3β/β-catenin pathway. For this study, elevated plus maze test was conducted for the determination of anxiety level. Immunohistochemistry for the expression of 5-HT and TPH expression and western blot for the expression of GSK3β and β-catenin were performed. We compared the effect of treadmill running on anxiety with fluoxetine.
MATERIALS AND METHODS
Experimental animals
The pregnant female Sprague-Dawley rats, weighing 280±10 g (10 weeks old), were used, and the day of delivery was designated postnatal day 0. This experiment study was approved by the Institutional Care and Use Committee of Kyung Hee University (KHUASP[SE]-17-099). On the postnatal day 14, the off spring randomly divided into 5 groups (n=8 in each group): the control group, the exercise group, the maternal separation group, the maternal separation and exercise group, and the maternal separation and fluoxetine-treated group.
Treadmill exercise protocol and treatment of fluoxetine
The rat pups in the exercise groups were forced to run on motorized treadmill (Columbus Instruments, Columbus, OH, USA) for 30 min once a day for 10 days, stating on the postnatal day 21. Exercise load consisted of treadmill at a speed 2 m/min for the first 30 min, without inclination. For the rat pups in the fluoxetine-treated group, 200 mL of fluoxetine (5 mg/kg; Sigma Chemical Co. St. Louis, MO, USA) was orally administrated once a day for 10 consecutive days, starting on the postnatal day 21.
Elevated plus maze test
The elevated plus maze test was performed on the postnatal day 30, according to a previously described method (Ko et al., 2013). The plus maze consisted of black acrylic with two open arms (50 cm×10 cm×36 cm) and two closed arms (50 cm×10 cm×36 cm), arranged so that the two arms were opposite and connected by a central platform (10 cm×10 cm). The whole plus maze was elevated 60 cm above the floor and illuminated by a 100-W light bulb fixed 2 m above the maze floor. During the test, each rat pup was placed on the central platform of the maze with their head facing an open arm. The latency time and number of entry in the open arms were collected during 7 min.
Tissue preparation
The animals were sacrificed after last treadmill exercise. After anesthetizing with Zoletil 50 (10 mg/kg intraperitoneally; Vibac Laboratories, Carros, France), 50 mM phosphate-buffered saline perfused to the rat pups, and then the rat pups were subsequently fixed with freshly prepared 500 mM phosphate buffer (pH, 7.4) containing 4% paraformaldehyde. The brains of the rat pups were removed and fixed in the same fixative overnight, and then transferred into a 30% sucrose solution for cryoprotection. Serial coronal sections with 40 μm thickness were obtained using a freezing microtome (Leica, Nussloch, Germany).
Immunohistochemistry
Immunohistochemistry was conducted to evaluate the 5-HT and TPH expression in the dorsal raphe, according to the previously described method (Ji et al., 2017; Moon et al., 2018). Free-floating tissue sections were incubated overnight with mouse anti-TPH or rabbit anti-5-HT antibody (1:1,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA). The sections were treated with biotinylated, anti-mouse TPH secondary antibody or with biotinylated anti-rabbit 5-HT secondary antibody (1:200; Vector Laboratories, Burlingame, CA, USA) during 1 hr. Next, avidin-biotin-peroxidase complex (Vector Laboratories) was treated to the section during 1 hour at 25°C. The sections were treated with a reaction mixture consisting of 0.03% 3,3′-diaminobenzidine and 0.03% hydrogen peroxide during 5 min. After the slides were dried under the room conditions, Permount (Fisher Scientific, Fair Lawn, NJ, USA) was used for the coverslips mounting.
Western blot analysis
Western blot analysis was performed, according to the previous method (Cho et al., 2018; Wu et al., 2015). The right hemisphere was homogenized on ice, and lysed in a lysis buffer containing 50 mM HEPES (pH 7.5), 150 mM NaCl, 10% glycerol, 1% Triton X-100, 1 mM PMSF, 1 mM EGTA, 1.5 mM MgCl2·6H2O, 1 mM sodium orthovanadate, and 100 mM sodium fluoride. Protein content was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein samples (30 μg) were separated on sodium dodecyl sulfate-polyacrylamide gel and transferred onto a nitrocellulose membrane. Mouse GSK3β antibody, mouse phosphorylated GSK3β (p-GSK3β) antibody, mouse β-catenin antibody, and mouse phosphorylated β-catenin (p-β-catenin) antibody (1:1,000; Santa Cruz Biotechnology) were used as the primary antibodies. Horseradish peroxidase-conjugated anti-mouse antibody (1:2,000; Vector Laboratories) for GSK3β, p-GSK3β, β-catenin, and p-β-catenin were used as secondary antibodies. Using a cold pack and prechilled buffer, membrane transfer was conducted at 4°C. Enhanced chemiluminescence detection kit (Santa Cruz Biotechnology) was used for band detection.
Data analysis
The numbers of 5-HT and TPH-positive cells in the dorsal raphe were counted hemilaterally through a light microscope (Olympus, Tokyo, Japan). The area of the dorsal raphe was measured using an Image-Pro plus computer-assisted image analysis system (Media Cybernetics Inc., Silver Spring, MD, USA) attached to a light microscope (Olympus). Molecular Analyst (ver. 1.4.1, Bio-Rad) was used for analyzing of detected bands.
Differences among the groups were evaluated using SPSS Statistics ver. 21.0 (IBM Co., Armonk, NY, USA) by the one-way analysis of variance followed by Duncan post hoc test. All values are expressed as the mean±standard error of the mean. Statistically significant differences were established at P<0.05.
RESULTS
Anxiety-like behavior
Anxiety-like behavior is presented in Fig. 1. Maternal separation decreased the number of entry and the time of latency in the open arms (P<0.05). Treadmill exercise and fluoxetine treatment increased the number of entry and the time of latency in the maternal separation rat pups (P<0.05).

Anxiety-like behavior in the elevated plus maze test. Left panel: percentage of open arms entries. Right panel: latency of open arms. A, control group; B, exercise group; C, maternal separation group; D, maternal separation and exercise group; E, maternal separation and fluoxetine-treated group. *P<0.05 compared to the control group. #P<0.05 compared to the maternal separation group.
Expression of 5-HT and TPH in the dorsal raphe
Expression of 5-HT and TPH in the dorsal raphe is presented in Fig. 2. Maternal separation decreased the number of 5-HT-positive and TPH-positive cells in the dorsal raphe (P<0.05). Treadmill exercise and fluoxetine treatment increased the number of 5-HT-positive and TPH-positive cells in the maternal separation rat pups (P<0.05).

5-Hydroxytryptamine (5-HT) and tryptophan hydroxylase (TPH) expression in the dorsal raphe. Left panel: photomicrographs of 5-HT-positive cells. Right panel: photomicrographs of TPH-positive cells. The scale bar represents 200 μm. □: Counting areas of the dorsal raphe. A, control group; B, exercise group; C, maternal separation group; D, maternal separation and exercise group; E, maternal separation and fluoxetine-treated group. *P<0.05 compared to the control group. #P<0.05 compared to the maternal separation group.
Expression of t-GSK3β and p-GSK3β in the hippocampus
Expression of t-GSK3β and p-GSK3β is presented in Fig. 3. Expression of t-GSK3β was not changed by maternal separation. However, maternal separation decreased the expression of p-GSK3β in the hippocampus (P<0.05). Treadmill exercise and fluoxetine treatment increased the expression of p-GSK3β in the maternal separation rat pups (P<0.05).
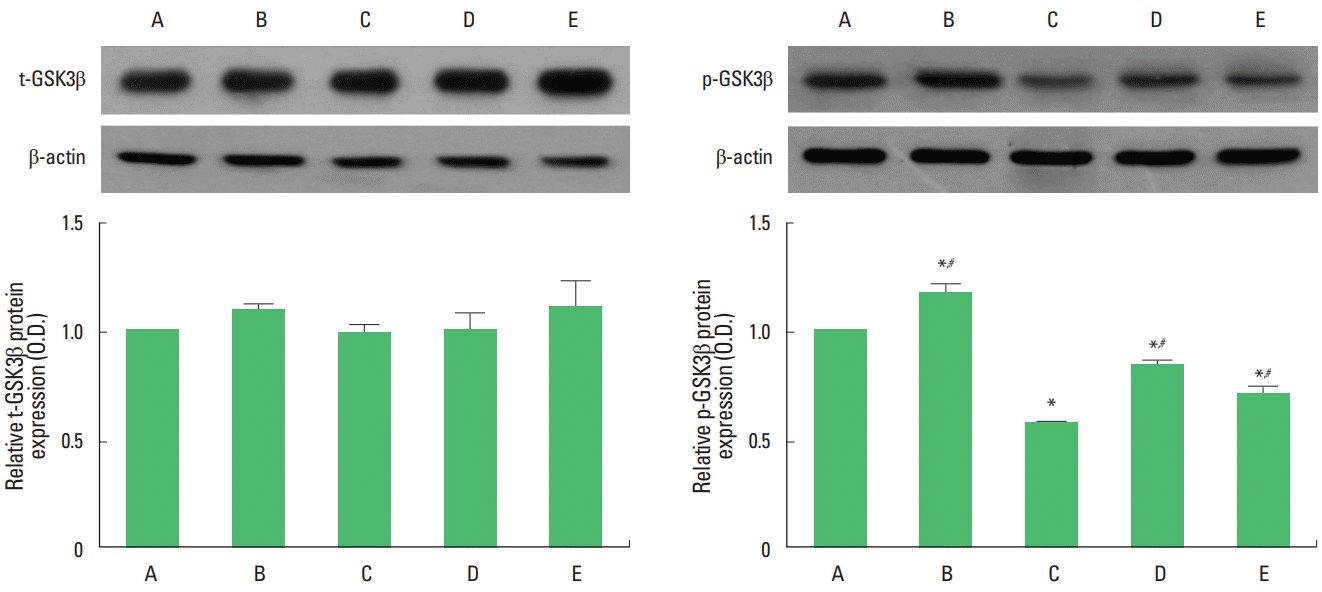
Effect of treadmill exercise on total glycogen synthase kinase 3 beta (t-GSK3β) and phosphorylated GSK3β (p-GSK3β) expression in the hippocampus. Left panel: representative expression of t-GSK3β in the hippocampus. Right panel: representative expression of p-GSK3β in the hippocampus. A, control group; B, exercise group; C, maternal separation group; D, maternal separation and exercise group; E, maternal separation and fluoxetine-treated group. *P<0.05 compared to the control group. #P<0.05 compared to the maternal separation group.
Expression of t-β-catenin and p-β-catenin in the hippocampus
The expression of t-β-catenin and p-β-catenin is presented in Fig. 4. Expression of t-β- was decreased by maternal separation (P<0.05). Treadmill exercise and fluoxetine treatment increased t-β-catenin expression in the maternal separation rat pups (P<0.05). Expression of p-β-catenin was increased by maternal separation (P<0.05). Treadmill exercise and fluoxetine treatment suppressed p-β-catenin expression in the maternal separation rat pups.

Total β-catenin (t-β-catenin) and phosphorylated β-catenin (p-β-catenin) expression in the hippocampus. Left panel: representative expression of t-β-catenin. Right panel: representative expression of p-β-catenin. A, control group; B, exercise group; C, maternal separation group; D, maternal separation and exercise group; E, maternal separation and fluoxetine-treated group. *P<0.05 compared to the control group. #P<0.05 compared to the maternal separation group.
DISCUSSION
Exercise is known to ameliorate depressive symptoms induced by maternal separation (Baek et al., 2012; Wu et al., 2017). The elevated plus maze is a test for the determination of anxiety level (Ko et al., 2013). In the present study, shorter latency time and less open arms entries were observed in the maternal separation rat pups, showing that maternal separation increased anxiety level. However, treadmill exercise and fluoxetine treatment lengthened latency time and increased open arms entries, showing that treadmill running reduced anxiety level. Previous studies suggested that exercise ameliorates anxiety-like behavior (Ko et al., 2013; Motaghinejad et al., 2017).
Depression or anxiety is associated with imbalance of neurotransmitters, such as serotonin, norepinephrine, and dopamine in the brain. Among them, 5-HT is closely related with the depression and anxiety (Baek et al., 2012; Ji et al., 2017; Motaghinejad et al., 2017). TPH is an enzyme that regulates 5-HT synthesis, and serotonin impairment in the suicide might be due to the hypofunction of serotonin synthesizing enzyme (Boldrini et al., 2005). In the present study, expression of 5-HT and TPH in the dorsal raphe was decreased by induction of maternal separation. However, treadmill exercise and fluoxetine treatment increased the expression of 5-HT and TPH in the dorsal raphe, suggesting that treadmill exercise and fluoxetine treatment restored the maternal separation-induced decrease of 5-HT and TPH expression. Our results confirmed that treadmill exercise and fluoxetine treatment alleviated maternal separation-induced anxiety status.
GSK3β inhibitors produce a weak anti-depressant-like effect and a strong anti-mania-like effect in bipolar disorder (Rowe et al., 2007). Inhibition of GSK3β in the hippocampus and striatum has anxiolytic and pro-social effect (Latapy et al., 2012). In the present study, t-GSK-β level in the hippocampus was not changed by maternal separation, however, the expression of p-GSK3β was decreased by maternal separation. The expression of 5-HT is closely related to the p-GSK3β expression. GSK3β is inhibited by SSRIs which used as antidepressants, and GSK3β is closely related to depression patients (Duman and Voleti, 2012; Inkster et al., 2009). In the hippocampus, fluoxetine regulates neurogenesis via up-regulating p-GSK3β, and this effect depends on 5-HT1A receptor (Hui et al., 2014; Li et al., 2004). In our study, treadmill exercise and fluoxetine treatment increased the expression of p-GSK3β in the hippocampus, demonstrating that treadmill exercise and fluoxetine enhances GSK3β phosphorylation via up-regulating 5-HT level.
GSK3β phosphorylation inhibits β-catenin decomposition by proteasome (Hwang et al., 2016). Inhibition of GSK3β activates β-catenin (Sen et al., 2009), and regular exercise inhibits stress-induced p-β-catenin expression (Leem et al., 2018). In the present study, expression of t-β-catenin in the hippocampus was decreased by maternal separation, in contrast, treadmill exercise and fluoxetine treatment increased t-β-catenin expression. Expression of p-β-catenin was increased by maternal separation, meanwhile, treadmill exercise and fluoxetine treatment suppressed p-β-catenin. In our study, treadmill exercise and fluoxetine regulate β-catenin activation via down-regulating β-catenin phosphorylation.
We demonstrated that treadmill exercise exerts alleviating effect on maternal separation-induced anxiety through enhancing 5-HT expression and GSK3β phosphorylation, and then inhibiting β-catenin phosphorylation. Treadmill exercise showed similar ameliorating effect on anxiety-like behavior as fluoxetine. Treadmill exercise acts as a useful strategy for the treatment of anxiety through GSK3β/β-catenin pathway.
Notes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.
ACKNOWLEDGMENTS
This work was supported by 2017 Sangmyung University Research Foundation of Korea.