Treadmill exercise improves depression-like symptoms by enhancing serotonergic function through upregulation of 5-HT1A expression in the olfactory bulbectomized rats
Article information
Abstract
The olfactory bulbectomy (OBX) is a well-known method inducing animal model of depression. Depression is associated with dysfunction of serotonin (5-hydroxytryptamine, 5-HT) system. In the present study, antidepressive effect of treadmill exercise was investigated using olfactory bulbectomized rats. After bilateral bulbectomy, the rats in the treadmill exercise groups were subjected to run on a treadmill for 30 min once a day during 28 days. Increased immobility time and decreased fast time in the forced swim test were observed in the olfactory bulbectomized rats. Sucrose preference in the sucrose preference test was decreased and activity in the open field test was also increased in the olfactory bulbectomized rats. Treadmill exercise decreased immobility time and activity and increased fast time and sucrose preference in the olfactory bulbectomized rats. Expressions of 5-HT and tryptophan hydroxylase (TPH) in the dorsal raphe of rats were suppressed by OBX and treadmill exercise increased the expressions of 5-HT and TPH in the olfactory bulbectomized rats. Serotonin receptor type 1A (5-HT1A) expression in the dorsal raphe was reduced by OBX and treadmill exercise increased 5-HT1A expression in the olfactory bulbectomized rats. In the present study, treadmill exercise ameliorated OBX-induced depressive symptoms. The antidepressive effect of treadmill exercise might be ascribed to the enhancement of serotonergic function through upregulation of 5-HT1A expression in the dorsal raphe.
INTRODUCTION
The olfactory bulbectomy (OBX) is a well-known method inducing animal model of depression (Douma et al., 2011; Sato et al., 2008). Removal of olfactory bulb caused behavioral, physiological, neurochemical, and morphological changes, similar to those observed in patients with depression (Douma et al., 2011). Increase of serotonergic dysfunction and pathologic findings of depression were observed in the olfactory bulbectomized rats (Sato et al., 2008).
Depression is associated with dysfunction of serotonin (5-hydroxytryptamine, 5-HT) system. 5-HT is closely related to the neuropsychiatric disorders, such as depression and anxiety (Arborelius 2004; Otsuka et al., 2016). Among 5-HT receptor subtypes, serotonin receptor type 1A (5-HT1A) and serotonin receptor type 2A (5-HT2A) are associated with the drugs for depression (Bhagwagar et al., 2006). 5-HT1A density was lower in the depressive rats (Sato et al., 2008). HT1A receptor in the hippocampus was deficit in the depression patients (Moses-Kolko et al., 2008). Tryptophan hydroxylase (TPH) catalyzes the rate-limiting step of serotonin biosynthesis in serotonin in the dorsal raphe (Rind et al., 2000).
Exercise has been used to treat various brain diseases. Stanton and Reaburn (2014) confirmed that exercise program exerted beneficial effect on depression. Regular physical exerted depressive symptoms (Archer et al., 2014). Treadmill exercise alleviated chronic mild stress-induced depressive symptoms in rats (Lee et al., 2015).
Many studies shown that exercise has a therapeutic effect on depression, however, the effect of treadmill exercise on OBX-induced depression is not clarified. In the present study, antidepressive effect of treadmill exercise was investigated using olfactory bulbectomized rats. For this study, forced swimming test (FST) for depressive symptoms, sucrose preference test for anhedonia, and open field test for activity were conducted. The serotonergic function in the dorsal raphe was detected using immunohistochemistry for 5-HT and TPH and Western blot for TPH and 5-HT1A.
MATERIALS AND METHODS
Animals
Female Sprague-Dawley rats (210±10 g, 6 weeks old) were used and the experimental procedures were performed in accordance with the animal care guidelines of the National Institutes of Health and the Korean Academy of Medical Sciences. The animals were housed under the controlled temperature (23°C±2°C) and lighting (08:00 a.m. to 20:00 p.m.) conditions with food and water available ad libitum. The animals were randomly divided into four groups (n=10 in each group): sham-operation group, sham-operation and treadmill exercise group, olfactory bulbectomized group, olfactory bulbectomized and treadmill exercise group.
Bilateral OBX surgery
OBX was performed using a previously described procedure (Kalshetti et al., 2015). The rats were anesthetized with isoflurane (2% isoflurane in 30% O2 and 70% N2; Vibac Laboratories, Carros, France). Each rat was placed in stereotaxic frame (Benchmark Deluxe, MyNeurolab, St. Louis, MO, USA), head was shaven, midline scalp sagittal incision (1 cm) was made, bilateral burr holes (diameter, 2 mm) was drilled at 8 mm anterior to bregma and 2 mm lateral from midline. Both main and accessory olfactory bulbs were aspirated through both burr holes using a blunt hypodermic needle attached to water pump. The rats in the sham-operation groups received operation for burr holes only. The rats were allowed to recover for the period of 7 days after surgery.
Treadmill exercise protocol
After completion of 7 days of recovery period, the rats in the treadmill exercise groups were subjected to run on a treadmill for 30 min once a day during 28 days. Exercise load consisted of running at a speed of 2 m/min for the first 5 min, at a speed of 5 m/min for the next 5 min, and then at a speed of 8 m/min for the last 20 min, with the 0° inclination.
Forced swimming test
In order to evaluate the degree of a depression state in rats, modified FST was conducted, as a previously described method (Baek et al., 2012). After the cessation of exercise, the rats conducted a pretest for 15 min to eliminate the acute stress by water and to adapt the animals to the water. Twenty-four hr after the pretest, the rats were tested for 6 min. The animals were placed individually into the glass cylinder 20 cm in diameter and 90 cm in height, filled with water to a height of 75 cm. The temperature of the water was adjusted to 25°C±1°C. During the test session, immobility time and fast time were analyzed using a Smart version 2.5 video tracking system (Panlab, Barcelona, Spain). Immobility was defined to occur when no additional activity was observed other than the actions needed to keep the rat’s head above the water. Fast was defined to occur when the rat was in an active vertical motion with its forelegs above the water level.
Sucrose preference test
Sucrose preference test for the evaluation of anhedonia was conducted, as a previously described method (Lee et al., 2015). For this purpose, two bottles were presented to the animals for 24 hr. One bottle contained pure drinking water and the other contained 1% sucrose solution. The sucrose preference (%) was calculated as follows: consumed total amount of sucrose/consume total amount of fluid (water+sucrose)×100 during 24 hr.
Open field test
To determine the activity in the rats, open-field test was conducted, as a previously described method (Roh et al., 2016). The rats were randomly assigned to an order of testing and placed in a white square open-field arena (100 cm×100 cm) made of wood, enclosed by 40-cm-high walls and exposed to strong illumination (200 lux). The arena was divided into 25 squares (20 cm×20 cm), consisting of 9 central and 16 peripheral squares. The animals were placed in the center of the arena and were allowed to explore the environment for 1 min. Following this time, the number of squares crossed was recorded for 5 min.
Tissue preparation
After the behavioral tests, the rats were deeply anesthetized with Zoletil 50 anesthesia (1 mg/kg, intraperitoneally; Vibac Laboratories). After complete anesthesia, the mice were transcardially perfused with 0.05-M phosphate-buffered saline (PBS), followed by 4% paraformaldehyde in 0.5-M sodium phosphate buffer (PB) at pH 7.4. The brain was removed, postfixed in the same fixative overnight, and transferred to a 30% sucrose solution for cryoprotection. Serial 40-μm-thick coronal sections were cut with a freezing microtome (Leica, Nussloch, Germany).
Immunohistochemistry for 5-HT and TPH expressions
Immunohistochemistry was performed for the detection of the 5-HT-positive and TPH-positive cells in the dorsal raphe, as a previously described method (Roh et al., 2016). Average eight sections were selected in each brain. The sections were incubated in PBS for 10 min and they were next washed 3 times with PBS. The sections were then incubated in 1% H2O2 for 30 min, and then they were incubated overnight with rabbit anti-5-HT antibody (Oncogene Research Product, Cambridge, UK) at a dilution of 1:500 for 5-HT expression and with mouse anti-TPH antibody (Oncogene Research Product) at a dilution of 1:500 for TPH expression. The sections were incubated for 1 hr with biotinylated anti-rabbit secondary antibody and with biotinylated anti-mouse secondary antibody (Vector Laboratories, Burlingame, CA, USA), and they were subsequently incubated with avidin-biotin-peroxidase complex (Vector Laboratories) for 1 hr. Immunoreactivity was visualized by incubating the sections in a solution consisting of 0.05% 3,3′-diaminobenzidine and 0.01% H2O2 in 50-mM Tris-buffer (pH, 7.6) for approximately 3 min. The sections were finally mounted on gelatin-coated glass slides. The slides were air-dried overnight at room temperature, and the coverslips were mounted using Permount (Thermo Fisher Scientific Inc., Waltham, MA, USA).
Western blot analysis for TPH and 5-HT1A expressions
Western blot was performed for the detection of the TPH and 5-HT1A expressions, as a previously described method (Kim et al., 2015). Sample tissues were lysed in ice-cold lysate buffer containing 50-mM HEPES (pH, 7.5), 150-mM NaCl, 10% glycerol, 1% Triton X-100, 1.5-mM magnesium chloride hexahydrate, 1-mM ethyleneglycol-bis-(β-aminoethyl ether)-N,N′-tetraacetic acid, 1-mM phenylmethylsulfonyl fluoride, 2-μg/mL leupeptin, 1-μg/mL pepstatin, 1-mM sodium orthovanadate, and 100-mM sodium fluoride, after which the mixture was incubated for 30 min at 4°C. The tissue debris was then removed by microcentrifugation followed by quick freezing of the supernatant. The protein concentration was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein 40 μg was separated on sodium dodecyl sulfate-polyacrylamide gels and transferred onto a nitrocellulose membrane (Whatman, Clifton, NJ, USA). Mouse anti-TPH antibody (1:1,000; Oncogene Research Product) and rabbit anti-5-HT1A antibody (1:1,000; Abcam, Cambridge, UK) were used as the primary antibodies. Horseradish peroxidase-conjugated anti-mouse antibody (1:3,000; Vector Laboratories) and horseradish peroxidase-conjugated anti-rabbit antibody (1:3,000; Vector Laboratories) were used as the secondary antibodies to probe TPH and 5-HT1A. Band detection was performed using the enhanced chemiluminescence detection system (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The bands were quantified using an Image-Pro Plus computer-assisted image analysis system (Media Cybernetics Inc., Bethesda, MD, USA).
Statistical analysis
Statistical analysis was performed using one-way analysis of variance followed by Duncan post hoc test, and the results were expressed as the mean±standard error of the mean. Significance was set as P<0.05.
RESULTS
Effect of treadmill exercise on immobility and fasting time
The immobility time in the olfactory bulbectomized group was longer than that in the sham-operation group. Treadmill exercise decreased the immobility time in the rats of the olfactory bulbectomized group (Fig. 1A). The fasting time in the olfactory bulbectomized group was shorter than that in the sham-operation group. Treadmill exercise increased the fasting time in the rats of the olfactory bulbectomized group (Fig. 1B).

Effect of treadmill exercise on immobility (A) and fast (B) time. Sham, sham-operation group; Sham-Ex, sham-operation and treadmill exercise group; OBX, olfactory bulbectomy group; OBX-Ex, olfactory bulbectomy and treadmill exercise group. Data are represented as the mean±standard error of the mean. *P<0.05 compared to the Sham group. #P<0.05 compared to the OBX group.
Effect of treadmill exercise on sucrose preference
The sucrose preference in the olfactory bulbectomized group was lower than that in the sham-operation group. Treadmill exercise increased the sucrose preference in the rats of the olfactory bulbectomized group (Fig. 2).

Effect of treadmill exercise on sucrose intake. Sham, sham-operation group; Sham-Ex, sham-operation and treadmill exercise group; OBX, olfactory bulbectomy group; OBX-Ex, olfactory bulbectomy and treadmill exercise group. Data are represented as the mean±standard error of the mean. *P<0.05 compared to the Sham group. #P<0.05 compared to the OBX group.
Effect of treadmill exercise on activity
The activity in the olfactory bulbectomized group was higher than that in the sham-operation group. Treadmill exercise decreased the activity in the rats of the olfactory bulbectomized group (Fig. 3).

Effect of treadmill exercise on activity. Sham, sham-operation group; Sham-Ex, sham-operation and treadmill exercise group; OBX, olfactory bulbectomy group; OBX-Ex, olfactory bulbectomy and treadmill exercise group. Data are represented as the mean±standard error of the mean. *P<0.05 compared to the Sham group. #P<0.05 compared to the OBX group.
Effect of treadmill exercise on 5-HT and TPH expressions in the dorsal raphe
The number of 5-HT-positive cells in the dorsal raphe was suppressed by OBX and the number of 5-HT-positive cells in the olfactory bulbectomized rats was recovered near to the normal level by treadmill exercise (Fig. 4A). The number of TPH-positive cells in the dorsal raphe was suppressed by OBX and the number of TPH-positive cells in the olfactory bulbectomized rats was recovered near to the normal level by treadmill exercise (Fig. 4B).
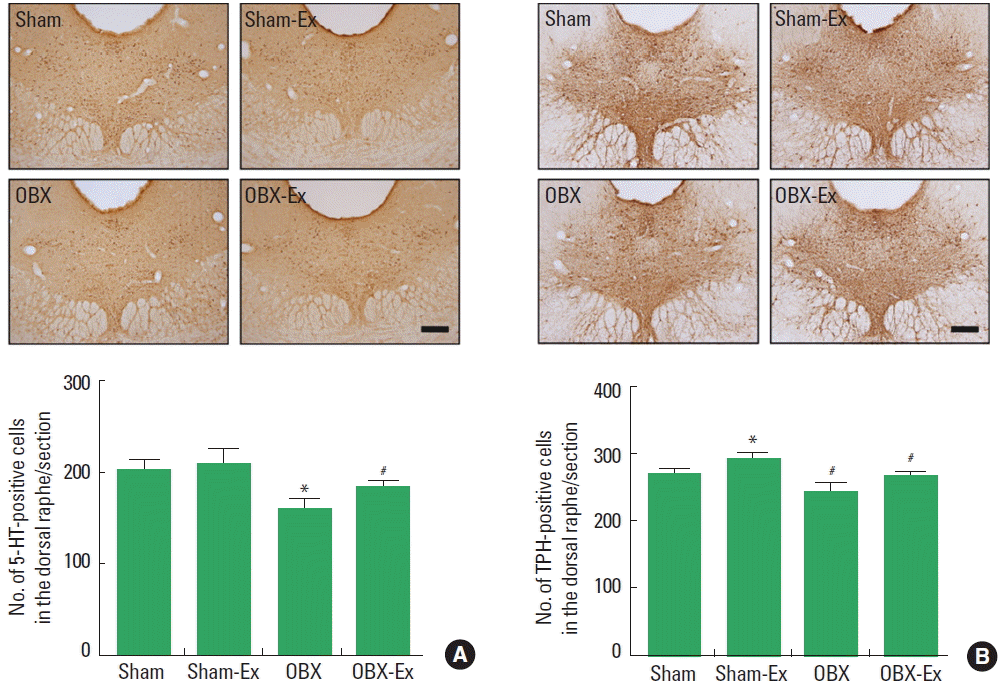
Effect of treadmill exercise on the numbers of 5-hydroxytryptamine (5-HT)-positive cells (A) and tryptophan hydroxylase (TPH)-positive cells (B) in the dorsal raphe. Upper panels: Photomicrographs of immunohistochemistry for the 5-HT-positive cells and the TPH-positive cells. Sham, sham-operation group; Sham-Ex, sham-operation and treadmill exercise group; OBX, olfactory bulbectomy group; OBX-Ex, olfactory bulbectomy and treadmill exercise group. Data are represented as the mean±standard error of the mean. *P<0.05 compared to the Sham group. #P<0.05 compared to the OBX group. The scale bar represents 250 μm.
Effect of treadmill exercise on TPH and 5-HT1A expressions in the dorsal raphe
The expression of TPH was decreased in the olfactory bulbectomized group but treadmill exercise increased the TPH expression in the rats of the olfactory bulbectomized group (Fig. 5A). The expression of 5-HT1A was decreased in the olfactory bulbectomized group but treadmill exercise increased the 5-HTA expression in the rats of the olfactory bulbectomized group (Fig. 5B).

Effect of treadmill exercise on tryptophan hydroxylase (TPH) protein (B) and 5-hydroxytryptamine 1A receptor (5-HT1A) protein (C) expressions in the dorsal raphe. (A) Western blot analysis. Sham, sham-operation group; Sham-Ex, sham-operation and treadmill exercise group; OBX, olfactory bulbectomy group; OBX-Ex, olfactory bulbectomy and treadmill exercise group. Data are represented as the mean±standard error of the mean. *P<0.05 compared to the Sham group. #P<0.05 compared to the OBX group.
DISCUSSION
The OBS rat model is widely accepted animal model of depression, showing serotonergic imbalance in the brain (Sato et al., 2008). Chronic stress procedure resulted in a strong decrease in sucrose preference, a putative indicator of anhedonia in rodents (Strekalova et al., 2004). OBX model is associated with increment of immobility time in the forced swimming test and anhedonic-like response in sucrose preference (Rinwa and Kumar, 2014). Olfactory bulbectomized rats demonstrated depressive-like behavioral marks, such as the decreased sucrose consumption, hyperactivity, impaired short-term memory, and anxiety-like behavioral features (Stepanichev et al., 2016). Treadmill exercise restored sucrose consumption with increased cell proliferation and decreased apoptotic cell death in the depressive rats (Lee et al., 2015).
In the present study, increased immobility time and decreased fast time in the forced swim test were observed in the olfactory bulbectomized rats, showing depressive symptoms. Sucrose preference in the sucrose preference test was decreased in the olfactory bulbectomized rats, showing anhedonic-like response. Activity in the open field test was also increased in the olfactory bulbectomized rats, showing depressive symptoms. Treadmill exercise decreased immobility time and activity and increased fast time and sucrose preference in the olfactory bulbectomized rats. The present results represent that treadmill exercise ameliorated depressive symptoms in the olfactory bulbectomized rats.
Suppression of the brain serotonergic system is implicated in the pathophysiology of depression (Arborelius et al., 2004). The symptoms of depression are closely associated with reduced activity of the serotonergic system (van der Stelt et al., 2005). The dorsal raphe is involved in the anxiolytic effect of physical activity (Greenwood et al., 2003). Treadmill exercise increased expressions of 5-HT and TPH in the dorsal raphe of the maternal separation-induced depressive rat pups (Baek et al., 2012) and of the chronic mild stress-induced depressive rats (Lee et al., 2015). Physical exercise may exert its antidepressive and anxiolytic effects through neuronal activation of 5-HT neurons in the dorsal raphe (Otsuka et al., 2016).
In the present study, 5-HT and TPH expressions in the dorsal raphe of rats were suppressed by OBX and treadmill exercise increased the expressions of 5-HT and TPH in the olfactory bulbectomized rats. The present results demonstrate that treadmill exercise restored serotonin synthesis in the dorsal raphe of olfactory bulbectomized rats.
Wheel running for 6 weeks increased 5-HT1A inhibitory autoreceptor mRNA expression in the dorsal raphe, suggesting that wheel running prevented behavioral depression/learned helplessness and attenuated 5-HT neural activity in dorsal raphe during uncontrollable stress (Greenwood et al., 2003). 5-HT1A is involved in the pathophysiology of depression and other stress-related disorders (Kieran et al., 2010). Chronic mild stress reduced both mRNA and protein levels of 5-HT1A (Szewczyk et al., 2014). Treadmill exercise alleviated prenatal stress-induced anxiety through 5-HT1A activation in rats (Lee et al., 2016).
In the present study, 5-HT1A expression in the dorsal raphe was reduced by OBX and treadmill exercise increased 5-HT1A expression in the olfactory bulbectomized rats. The present results demonstrate that treadmill exercise up-regulated 5-HT1A expression in the dorsal raphe of olfactory bulbectomized rats.
In the present study, treadmill exercise ameliorated OBX-induced depressive symptoms. The antidepressive effect of treadmill exercise might be ascribed to the enhancement of serotonergic function through upregulation of 5-HT1A expression in the dorsal raphe. Based on the present results, treadmill exercise might be effective strategy for depression patients.
ACKNOWLEDGMENTS
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2014S1 A5A2A01015472).
Notes
CONFLICT OF INEREST
No potential conflict of interest relevant to this article was reported.