Antidepressants modulate glycine action in rat hippocampus
Article information
Abstract
Antidepressants are drugs that relieve symptoms of depressive disorders. Fluoxetine, tianeptine, and milnacipran are different types of antidepressants, and they have widely been used for relieving of depression symptoms. In the present study, the effects of fluoxetine, tianeptine, and milnacipran on the glycine-induced ion current by nystatin-perforated patch clamp and on the amplitude of field potential in the hippocampal CA1 region by multichannel extracellular recording, MED64, system, were studied. In the present results, fluoxetine, tianeptine, and milnacipran reduced glycine-induced ion current in the hippocampal CA1 neurons in nystatin-perforated patch clamp method. These drugs enhanced the amplitude of the field potential in the hippocampal CA1 region in MED64 system. These results suggest that antidepressants may increase neuronal activity by enhancing field potential through inhibition on glycine-induced ion current.
INTRODUCTION
Antidepressants are drugs that relieve symptoms of depressive disorders. There are above 30 different kinds of antidepressants available today, and these drugs are classified into four major groups: the monoamine oxidase inhibitors, the tricyclic antidepressants, the selective serotonin reuptake inhibitors (SSRIs), and the others called as heterocyclic antidepressants. Among them, SSRIs are widely used for the treatment of depression.
Fluoxetine, tianeptine, and milnacipran are different types of antidepressants, and they have widely been used for relieving of depression symptoms. Fluoxetine is a classic SSRI, and has been shown to be highly effective with little side effects (Bymaster et al., 2002). Tianeptine belongs to the selective serotonin reuptake enhancer (SSRE) (Fattaccini et al., 1990). Milnacipran is a serotonin and norepinephrine (NE) reuptake inhibitor (SNRI) (Delini-Stula, 2000). These antidepressants are related with serotonin (5-hydroxytryptamine, 5-HT) theory. Fluoxetine selectively inhibits serotonin reuptake pump from serotonergic nerve terminals, resulting in increment of serotonin concentration in synaptic cleft (Schmidt et al., 1988). Tianeptine increases the level of 5-hydroxyindoleacetic acid in hippocampus and hypothalamus, enhancing 5-HT uptake and its metabolism (Marinesco et al., 1996). Milnacipran acts on the 5-HT and NE neurons, and shows long-term efficacy in patients with major depression (Mongeau et al., 1998). Chronic administration of milnacipran significantly increases basal synthesis of 5-HT and NE (Kamata et al., 2003).
The amino acid glycine is a major inhibitory neurotransmitter in the brain and spinal cord. Glycine has two physiological synaptic functions: it enhances the N-methyl-D-aspartate response at concentrations less than 1 μM, and at higher concentrations, it activates strychnine-sensitive glycine receptors (GLYRs), resulting in chloride-mediated neuronal inhibition (Pourcho et al., 1992). GLYRs are widely distributed in the hippocampus besides spinal cord and brain stem (Chattipakorn and McMahon, 2002). Glycine synapses in the brain stem and spinal cord are implicated in the transmission of nociception, in which glycine inhibits neurotransmission and relieves pain. Moreover, some glycine transpoters are associated with neurotransmission by modulation of glycine concentration at the synaptic cleft in glycinergic neurons. Chronic treatment with antidepressants reduced the potency of glycine (Nowak et al., 1993).
Field potential is extracellular potential induced by membrane electromotive force. The field potential at any point in the medium is given by the sum of potentials resulting from each of the current sourced, weighed according to their distances and the distribution of conductivity in the medium (Zhou and Poon, 2000). Measuring of the field potential is a technique that has widely been used for the synaptic transmission in the hippocampus, neocortex, and other brain regions. Field potential in the hippocampal CA1 region represents field excitatory postsynaptic potential (fEPSP) (Zhou and Poon, 2000). Simultaneous recording of many sites in the brain enables the detection of brain networks in spatiotemporal patterns (Shimono et al., 2002).
The neurophysiological mechanisms underlying the antidepressants still remains unclear, and the exact mechanisms in relation with neuronal excitability and synaptic transmission in the hippocampus have not been clarified yet. In the present study, the effects of fluoxetine, tianeptine, and milnacipran on the glycine-induced ion current by nystatin-perforated patch clamp and on the amplitude of field potential in the hippocampal CA1 region by multichannel extracellular recording, MED64, system, were studied.
MATERIALS AND METHODS
Patch clamp recording
Preparation of hippocampal CA1 neurons
Hippocampal CA1 neurons were freshly dissociated using technique described previously elsewhere (Kim et al., 2002; Yang et al., 2003). In brief, 10- to 15-day-old Sprague-Dawley rats of both sexes were decapitated under Zoletil 50 anesthesia (50 mg/kg; intramuscular).
The brain was removed and the transverse slices (400-μm thickness) were made with a microslicer (DTK-1000, DSK, Tokyo, Japan). Slices were preincubated in the incubation solution that had been well saturated with 95% O2 and 5% CO2 at room temperature for 30 min. Then, the slices were treated with pronase (protease XIV, 1 mg/6 mL of the oxygenated incubation solution) for 40–80 min at 32°C and subsequently with thermolysin (protease X, 1 mg/6 mL) for 10–20 min at 32°C. After enzyme treatment, the slices were kept in the enzyme free incubation solution for 1 hr.
The hippocampal CA1 region was identified in a 60-mm culture dish coated with silicone under a binocular microscope (SZ-ST, Olympus, Tokyo, Japan), and was micropunched out from the slices with an electrolytically polished injection needle. The micropunched hippocampal CA1 neurons were mechanically dissociated in a different dish with fire-polished fine glass Pasteur pipettes in 35-mm plastic culture dishes (3801, Falcon, Franklin Lakes, NJ, USA) filled with standard solution. The dissociation procedure was done under an inverted phase-contrast microscope (CK-2, Olympus, Tokyo, Japan). The dissociated neurons usually adhered to the bottom of the dish within 20 min. These cells were remained viable for electrophysiological studies up to 6 hr after dissociation.
Electrical measurements
Electrical recordings were performed in the nystatin-perforated patch recording mode under voltage-clamp condition (Kim et al., 2002; Yang et al., 2003). Patch pipette was prepared from glass capillaries with an outer diameter of 1.5 mm on a two-stage puller (PB-7, Narishige, Tokyo, Japan). The resistance between the recording electrode filled with the internal pipette solution and the reference electrode was 6–8MΩ. After stable perforated patch formation, the series resistance ranged from 16 to 25MΩ.
Electrical stimulation, current recordings, and filtration of currents (at 2.9 kHz) were obtained with an EPC-7 patch-clamp amplifier (List-Electronic, Darmstadt/Eberstat, Germany). The current and voltage were monitored on a pen recorder (Recti-Horiz-8K, NEC San-ei, Tokyo, Japan). All experiments were performed at room temperature (22°C–24°C).
Drugs were added to the standard solution at the final concentrations provided in the text and were applied using a rapid application system termed the “Y-tube method” as described elsewhere (Kim et al., 2002; Yang et al., 2003). By this technique, the standard solution surrounding a neuron could be exchanged within 10–20 ms.
MED64 system recording
Preparation of hippocampal slices
The general procedure for the preparation of the hippocampal slices was same as previously described (Shimono et al., 2002; Yi et al., 2009). In brief, 3- to 4-week-old Sprague-Dawley rats of both sexes were sacrificed by decapitation after anesthesia using Zoletil 50 (50 mg/kg; intramuscular), and whole brain was removed carefully. The brain was immediately soaked in ice-cold oxygenated incubation solution within 1 min. Appropriate portions containing hippocampus of the brain were placed on the ice-cold plate of the vibrating tissue slicer (DTK-1000, DSK) and transverse slices (300-μm thickness) were made. Slices were incubated in the incubation solution which had been well saturated through exposure to 95% O2 and 5% CO2 for 1 hr at room temperature.
Electrical measurements
Electrophysiological recording was measured by 64-channel multielectrode dish of the MED64 system (Alpha MED Science, Tokyo, Japan), a novel two-dimensional electroactivity monitoring technique described elsewhere (Shimono et al., 2002; Yi et al., 2009). A slice was placed on the center on the MED probe, which was treated with coating solution for 8 hr at room temperature, and positioned to cover the 8×8 microelectrode arrays with 150-μm interpolar distance. After positioning of the slice on MED probe, microscopic photograph was taken. MED probe was placed on the MED connector and incubation solution oxygenated with 95% O2 and 5% CO2 was continuously infused at the rate of 2 mL/min (at 34°C). Evoked field potentials on 64 sites were simultaneously recorded after selecting of one recording site and two stimulation sites. For the recording of evoked field potentials, a pair of single planar microelectrodes with bipolar constant current pulses (45–90 μA, 0.1 ms) was applied. Amplitude of field potential recorded in the absence of drug was used as a control value, and then amplitude of field potential recorded during drug application was measured.
Solutions
The ionic composition of the incubation solutions was (in mM): NaCl 124, KCl 5, KH2PO4 1.2, MgSO4 1.3, CaCl2 2.4, glucose 10, and NaHCO3 24. The pH was adjusted to pH 7.4 by continuous bubbling with 95% O2 and 5% CO2 and was maintained at 34°C in water bath. The composition of the standard external solution was (in mM): NaCl 150, KCl 5, MgCl2 1, CaCl2 2, glucose 10, and N-2-hydroxyethylpiperazine-N′-2-ethanesulphonic acid (HEPES) 10. The pH was adjusted to the 7.4 with tris-hydroxymethylaminomethane (Tris-base). The composition of the internal pipette solution for nystatin perforated recording contained (in mM): KCl 150 and HEPES 10. The pH was adjusted to 7.2 by adding Tris-base. A stock solution containing 10 mg/mL nystatin in methanol was prepared and added in a final concentration of 200 μg/mL to the patch pipette solution. The composition of the probe coating solution contained 0.1% polyethylenimine in 25 mM borate buffer adjusted pH 8.4.
Drugs
Zoletil 50 was obtained from Vibac Laboratories (Carros, France). Fluoxetine was purchased from Sigma Chemical Co. (St. Louis, MO, USA). Tianeptine was obtained from SERVIER (Courbevoie, France). Milnacipran was obtained from Cypress Bioscience Inc. (San Diego, CA, USA). Glycine, polyethylenimine, pronase, thermolysin, nystatin, and most of the other drugs used in this study were purchased from Sigma Chemical Co.
Statistical analysis
The data was analyzed was performed using one-way analysis of variance followed by Tukey multiple comparison tests. The results were expressed as the mean±standard error of the mean and P-value less than 0.05 was considered as a statistical significance.
RESULTS
Effects of antidepressants on glycine-induced ion current in rat hippocampal CA1 neurons
In the nystatin-perforated patch-clamp mode, experiments were carried out at a holding potential (VH) of −50 mV. To investigate the modulation of antidepressants on the glycine-induced ion current, the magnitude of ion current elicited by 10−5 M glycine was used as the control value set as 1. Antidepressants were applied every 2 min and ion current-activated by 10−5 M glycine.
Modulation of fluoxetine on glycine-induced ion current
To investigate the modulation of fluoxetine on the glycine-induced ion current, the magnitude of ion current elicited by 10−5 M glycine was used as the control value and 10−7 M, 10−6 M, 10−5 M, and 10−4 M fluoxetine were applied simultaneously with 10−5 M glycine. Fluoxetine at concentrations of 10−7 M, 10−6 M, 10−5 M, and 10−4 M suppressed glycine-induced ion current to 0.99%±0.03%, 0.84%±0.06%, 0.77%±0.05%, and 0.66%±0.04%, with the control value set as 1. In the present study, the glycine-induced ion current in the hippocampal CA1 neurons was suppressed by application of fluoxetine (Fig. 1).

Modulation of fluoxetine on glycine-activated ion current. Glycine-induced ion current was significantly inhibited by 10−5 M, and 10−4 M of fluoxetine. *P<0.05 compared to the control.
Modulation of tianeptine on glycine-induced ion current
To investigate the modulation of tianeptine on the glycine-induced ion current, the magnitude of ion current elicited by 10−5 M glycine was used as the control value and 10−7 M, 10−6 M, 10−5 M, and 10−4 M tianeptine were applied simultaneously with 10−5 M glycine. Tianeptine at concentrations of 10−7 M, 10−6 M, 10−5 M, and 10−4 M suppressed glycine-induced ion current about 0.99%±0.03%, 0.84%±0.06%, 0.77%±0.07%, and 0.63%±0.03% of the control value set as 1. In the present study, the glycine-induced ion current in the hippocampal CA1 neurons was suppressed by application of tianeptine (Fig. 2).
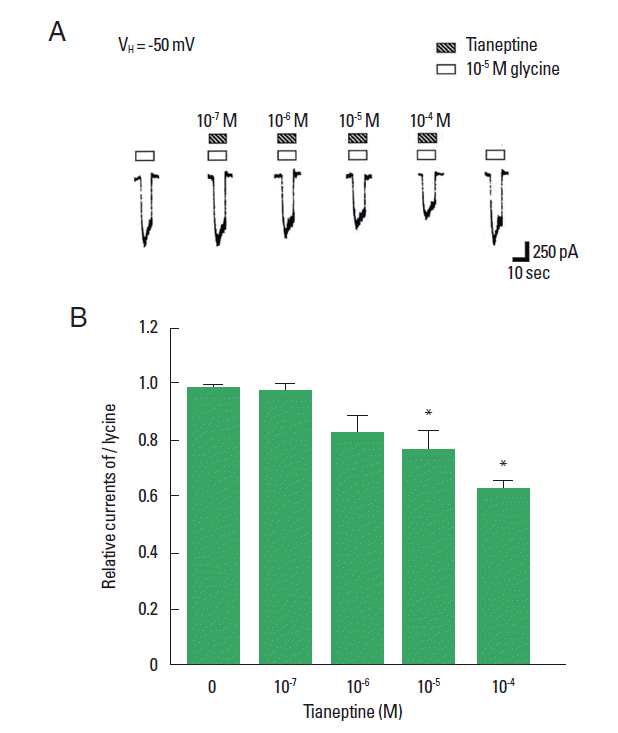
Modulation of tianeptine on glycine-activated ion current. Glycine-induced ion current was significantly inhibited by 10−5 M, and 10−4 M of tianeptine. *P<0.05 compared to the control.
Modulation of milnacipran on glycine-induced ion current
To investigate the modulation of milnacipran on the glycine-induced ion current, the magnitude of ion current elicited by 10−5 M glycine was used as the control value and 10−7 M, 10−6 M, 10−5 M, and 10−4 M milnacipran were applied simultaneously with 10−5 M glycine. Milnacipran at concentrations of 10−7 M, 10−6 M, 10−5 M, and 10−4 M suppressed glycine-induced ion current about 0.99%±0.08%, 0.86%±0.03%, 0.81%±0.04%, and 0.67%±0.05% of the control value set as 1. In the present study, the glycine-induced ion current in the hippocampal CA1 neurons was suppressed by application of milnacipran (Fig. 3).

Modulation of milnacipran on glycine-activated ion current. Glycine-induced ion current was significantly inhibited by 10−5 M, and 10−4 M of milnacipran. *P<0.05 compared to the control.
Effect of glycine on antidepressants-induced field potential in rat hippocampal slices
Experiment was carried out by applying a fixed pulse of 45–90 μV. Amplitude of the field potential in the CA1 region was recorded without drug applications for 10 min, and used as a control value. Each antidepressant was applied at the rate of 2 mL/min for 10 min following measuring of control value. The amplitude change induced by antidepressants was expressed as a percent (%) of the control value set as 100%.
Effect of glycine on fluoxetine-induced field potential
In order to evaluate the involvement of the inhibitory neurotransmitter glycine on fluoxetine-activated field potential in rat hippocampal slices, 10−5 M of glycine was coapplied on the hippocampal slices with 10−5 M of fluoxetine and amplitude of the field potential was recorded for 20 min. The amplitude of field potential in the CA1 region was similarly continued to 100.58%±2.93% of the control value set as 100% by treatment with 10−5 M of glycine and 10−5 M of fluoxetine. Subsequent treatment with 10−5 M fluoxetine in the absence of glycine for 20 min enhanced the field potential to 135.50%±6.20% of the control value. After maximum increase of field potential by fluoxetine treatment, hippocampal slices was washed with incubation solution for 40 min. The field potential enhanced by fluoxetine application was diminished to 114.50%± 8.20% of the control value by wash out (Fig. 4).
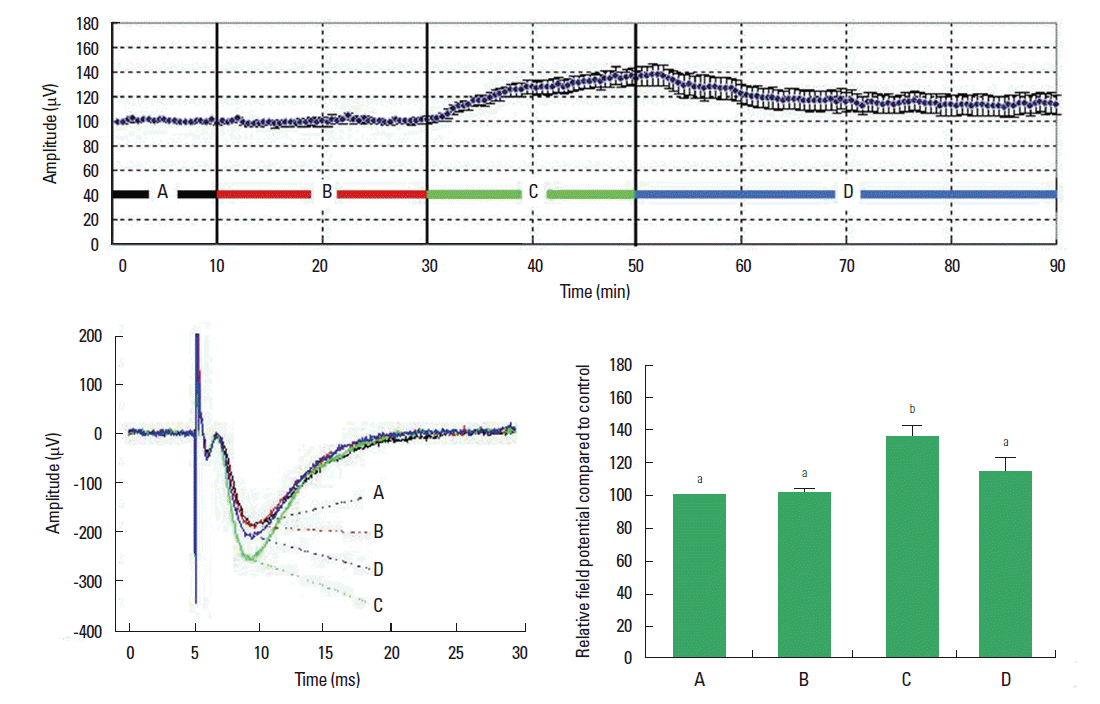
Effect of glycine on fluoxetine-induced field potential amplitude in rat hippocampal slices. Above: Dot plot of relative field potential amplitude with time. Each dot was recoded as 30-sec intervals. Left below: Schematic representation of field potential. Right below: Relative field potential compared to control. Letters (a, b) mean statistical significance P<0.05. (A) Control group in the absence of any drugs, (B) 10−5 M of glycine and 10−5 M fluoxetine-treated group, (C) 10−5 M fluoxetine-treated group, (D) washing group with incubation solution.
In the present study, coapplication of glycine and fluoxetine inhibited the field potential to the control value, but treatment with only fluoxetine was shown to enhance field potential amplitude in the CA1 region and of hippocampus. The increased field potential by fluoxetine application was recovered as control value by wash out.
Effect of glycine on tianeptine-induced field potential
In order to evaluate the involvement of the inhibitory neurotransmitter glycine on tianeptine-activated field potential in rat hippocampal slices, 10−5 M of glycine was coapplied on the hippocampal slices with 10−5 M of tianeptine and amplitude of the field potential was recorded for 20 min. The amplitude of field potential in the CA1 region was similarly continued to 110.98%±4.25% of the control value set as 100% by treatment with 10−5 M of glycine and 10−5 M of tianeptine. Subsequent treatment with 10−5 M tianeptine in the absence of glycine for 20 min enhanced the field potential to 157.76%±9.41% of the control value. After maximum increase of field potential by tianeptine treatment, hippocampal slices was washed with incubation solution for 40 min. The field potential enhanced by tianeptine application was diminished to 125.98%±14.07% of the control value by wash out (Fig. 5).

Effect of glycine on tianeptine-induced field potential amplitude in rat hippocampal slices. Above: Dot plot of relative field potential amplitude with time. Each dot was recoded as 30-sec intervals. Left below: Schematic representation of field potential. Right below: Relative field potential compared to control. Letters (a, b) mean statistical significance P<0.05. (A) Control group in the absence of any drugs, (B) 10−5 M of glycine and 10−5 M tianeptine-treated group, (C) 10−5 M tianeptine-treated group, (D) washing group with incubation solution.
In the present study, coapplication of glycine and tianeptine inhibited the field potential to the control value, but treatment with only tianeptine was shown to enhance field potential amplitude in the CA1 region and of hippocampus. The increased field potential by tianeptine application was partially recovered to control value by washout.
Effect of glycine on milnacipran-induced ion current
In order to evaluate the involvement of the inhibitory neurotransmitter glycine on milnacipran-activated field potential in rat hippocampal slices, 10−5 M of glycine was coapplied on the hippocampal slices with 10−5 M of milnacipran and amplitude of the field potential was recorded for 20 min. The amplitude of field potential in the CA1 region was similarly continued to 101.10%±3.44% of the control value set as 100% by treatment with 10−5 M of glycine and 10−5 M of milnacipran. Subsequently treatment with 10−5 M milnacipran in the absence of glycine for 20 min enhanced the field potential to 147.14%±7.60% of the control value. After maximum increase of field potential by milnacipran treatment, hippocampal slices was washed with incubation solution for 40 min. The field potential enhanced by milnacipran application was diminished to 106.49%±5.02% of the control value by wash out (Fig. 6).

Effect of glycine on milnacipran-induced field potential amplitude in rat hippocampal slices. Above: Dot plot of relative field potential amplitude with time. Each dot was recoded as 30-sec intervals. Left below: Schematic representation of field potential. Right below: Relative field potential compared to control. Letters (a, b) mean statistical significance P<0.05. (A) Control group in the absence of any drugs, (B) 10−5 M of glycine and 10−5 M milnacipran-treated groupa, (C) 10−5 M milnacipran-treated group, (D) washing group with incubation solution.
In the present study, coapplication of glycine and milnacipran inhibited the field potential to the control value, but treatment with only milnacipran was shown to enhance field potential amplitude in the CA1 region and of hippocampus. The increased field potential by milnacipran application was recovered as control value by wash out.
DISCUSSION
In the present study, we investigated the modulation of antidepressants on the glycine-induced ion current. The results showed that fluoxetine, tianeptine, and milnacipran reduced glycine-induced ion current in the hippocampal CA1 neurons (Figs. 1–3).
Glycine application to GLYRs expressing neurons in acutely dissociated hippocampal slices in rats induced strychnine-sensitive currents which suppress neuronal excitability (Chattipakorn and McMahon 2002). GLYRs are expressed throughout the hippocampus, and they are characterized as glycine-gated chloride current in rat hippocampal slices pharmacologically (Brackmann et al., 2004; Mori et al., 2002). Binding of glycine to its receptors produces a large increase in chloride conductance, which causes membrane hyperpolarization (Chen et al., 1996).
In the present study, fluoxetine, tianeptine, and milnacipran enhanced field potential in the hippocampal CA1 region, independently to the types of these antidepressants such as SSRI, SSRE, and SNRI (Figs. 4–6). Moreover the magnitudes of field potential amplitude were maintained control level by coapplication of glycine and antidepressants. These results suggest that antidepressants inhibited glycine action, thus enhanced field potential in the hippocampal CA1 region. However extrasupplementation of glycine overcame the inhibitory action of antidepressants, thus field potential amplitude was maintained at control level. Here in this study, we have shown that antidepressants have inhibitory effect on glycine action.
Field potential represents the net balance of all currents that flow into a small, local volume in the extracellular fluid (Zhou and Poon, 2000). Release of excitatory neurotransmitters causes a local change in membrane potential, named excitatory postsynaptic potential (EPSP). EPSP generates a potential gradient along the membrane that, in turn, gives rise to the field potential, which reflects activities in hundreds to thousands of neurons in the brain.
In some studies, antidepressants, for example trimipramine, were reported not to increase the population and amplitude of spike potential in the hippocampus or eventually reduced the population and amplitude (Langosch and Walden, 2002). In other studies, however, it was reported that fEPSP in the hippocampus was increased by antidepressants. The repeated administration of imipramine enhanced adenosine-induced fEPSP (Zahorodna and Bijak, 1999).
In the present results, fluoxetine, tianeptine, and milnacipran reduced glycine-induced ion current in the hippocampal CA1 neurons in nystatin-perforated patch clamp method. Fluoxetine, tianeptine, and milnacipran enhanced the amplitude of the field potential in the hippocampal CA1 region in multichannel extracellular recording MED64 system. The magnitudes of field potential amplitude were maintained control level by coapplication of glycine and antidepressants. These results show that antidepressants can increase neuronal activity by enhancing field potential through inhibition on glycine-induced ion current.
ACKNOWLEDGMENTS
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2010-0022895).
Notes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.