INTRODUCTION
Different variants of serotonergic and dopaminergic substances may modulate the influence of environmental conditions on a range of psychological conditions. Depending on their genotype, people may have different sensitivities to the environmental conditions they encounter (Keltikangas-Järvinen and Salo, 2009).
Decreased dopamine and increased serotonin (5-hydroxytryptamine) have been linked to depressed psychological condition (Soares et al., 2007). Dopamine and serotonin are the important monoamine neurotransmitters known to be modulated by physical activity (Lin and Kuo, 2013). Acute exhaustive endurance exercise increases the synthesis, metabolism, and concentration of serotonin in several brain regions. Therefore, serotonin may play a role in central fatigue after exhaustive and prolonged exercise (Blomstrand, 2001). For this reason, the central fatigue hypothesis suggests that increased concentrations of serotonin in the brain impair central nervous system (CNS) functioning during prolonged exercise, resulting in the deterioration of exercise performance (Newsholme et al., 1992). Though the role of serotonin on central fatigue is well known, it is likely that other neurotransmitters such as dopamine are capable of influencing fatigue. Animal studies proved that brain dopamine and serotonin activities were increased during exercise, however, a distinguished fall in tissue dopamine concentration was explicit at the point of exhaustion (Davis and Bailey, 1997).
Exercise can inhibit the synthesis of serotonin caused by increased concentrations of dopamine in some parts of the brain and thereby delay fatigue (Chaouloff et al., 1987). Therefore, the ratio of serotonin to dopamine activity may be a major indicator linked to central fatigue. Also, prolactin may be a competent measure of the relationship between dopamine and serotonin (Freeman et al., 2000) as well as urgent fatigue during not compensable heat stress. Administration of caffeine was found to increase dopamine release in animal studies (Okada et al., 1996; Solinas et al., 2002) and also has the potential to reduce serotonin levels during exercise (Lim et al., 2001). Thermal stress can depress mood and motivation (Meeusen et al., 2006) is associated with thermal stress, implying a potential relationship between serotonin and dopamine.
We have been studying about physiological responses such as body temperature, sweating and endocrine responses during exercise or body immersion in hot water in relation to caffeine ingestion. (Bae et al., 2006; Lee and Kim, 2014; Lee et al., 2013). However, this is the first research to compare the alteration in serum serotonin and dopamine after acute caffeine ingestion via half immersion in hot water. The results of the “serotonin-fatigue hypothesis” from animal experiments are clear, however, are not clear on humans. Therefore, this study’s aim is to prove the effect of caffeine on serotonin and dopamine levels during and after passive heat loading in humans.
MATERIALS AND METHODS
Subjects
Thirty male volunteers participated and the physical characteristics of the subjects are summarized in Table 1. None of the subjects ingested caffeine habitually and none were acclimated or acclimatization. Inclusion criteria were no side effects from caffeine, no health problems, and no history of smoking. Each subject returned written informed consent to participate in the study, after being thoroughly acquainted with the purpose and the experimental procedures, as well as any potential risks. Subjects fasted for 6 hr and had to drink ~5–7 mL/kg of tap water 4 hr before the test on the day of the test. They were instructed to refrain from alcohol consumption and medications 24 hr before the test. All experimental protocols were approved by the Soonchunhyang University Research Committee.
Measurement and experimental procedure
The experiment was randomized. The tests were performed in a climate chamber from 2–5 p.m. We conducted this experiment from 2–5 p.m. to control for the influence of the body temperature circadian rhythm (Lee and Kim, 2014; Lee et al., 2013). After the subjects arrived at the laboratory, urine specific gravity was tested with a urine strip (Uriscan, Seoul, Korea) to confirm hydration equilibrium and blood samples were collected. The subjects were then randomly divided into 2 groups: group A (n=15) drank 200 mL of tap water (CON) while group B (n=15) drank 200 mL of tap water and 3-mg/kg caffeine (CAFF). All subjects sat in a chair in a relaxed posture for 60 min. Passive heat loading was conducted for 30 min with no drinking. Blood was sampled at −60 min before passive heat loading (Pre) and immediately after passive heat loading for 30 min (Post). After 60 min of resting (Post-60), blood sampling was performed again.
Passive heat loading
Passive heat loading was conducted in the city of Cheonan, the Republic of Korea. Cheonan is located in the south western part of Korea (126°52′N, 33.38′E) and extends northeast (130°4′N, 43.0′E). The environmental conditions of the test room were maintained at 24.5°C±0.3°C, 50%±3.0% relative humidity, and 1-m/sec air velocity. Half immersion in 42°C hot water was applied for 30 min. Previous studies have used a range of similar thermal intensity and time (Lee and Kim, 2014; Lee et al., 2013). Half immersion was applied up to the umbilicus in hot water.
Serotonin, dopamine, and prolactin level analysis
Blood samples were collected at Pre, Post, and Post-60 time points. Blood samples were collected in serum-separating tube from the antecubital vein and kept at room temperature for 15–30 min to allow for clotting. Afterward, the samples were centrifuged at 3,000 rpm (2,000×g) and 4°C for 10 min. The serum was then harvested and stored at −80°C until analysis. Serotonin level was determined using the high-pressure liquid chromatography (HPLC) method (Serotonin Kit, Recipe, Munich, Germany) with Alliance Waters 465 HPLC (Waters, Milford, MA, USA). Prolactin level was determined using the electrochemiluminescence immunoassay method (Prolactin II reagent, Roche, Mannheim, Germany) with Modular Analytics E (Roche, Mannheim, Germany). Plasma was then extracted and stored at −80°C until analysis. The dopamine level was determined using the HPLC method (Plasma catecholamine Kit, Bio-Rad, Munich, Germany) with Acclaim HPLC (Bio-Rad, Hercules, CA, USA).
Data analyses
Descriptive statistics are expressed as mean±standard deviation using commercially available computer software (IBM SPSS Statistics ver. 20.0, IBM Co., Armonk, NY, USA). Statistical significance was assessed by a two-way measure analysis of variance. Wilcoxon signed-rank test or Mann Whitney U-test were used to examine differences between the groups and within groups. Significant differences were assumed at P<0.05.
RESULTS
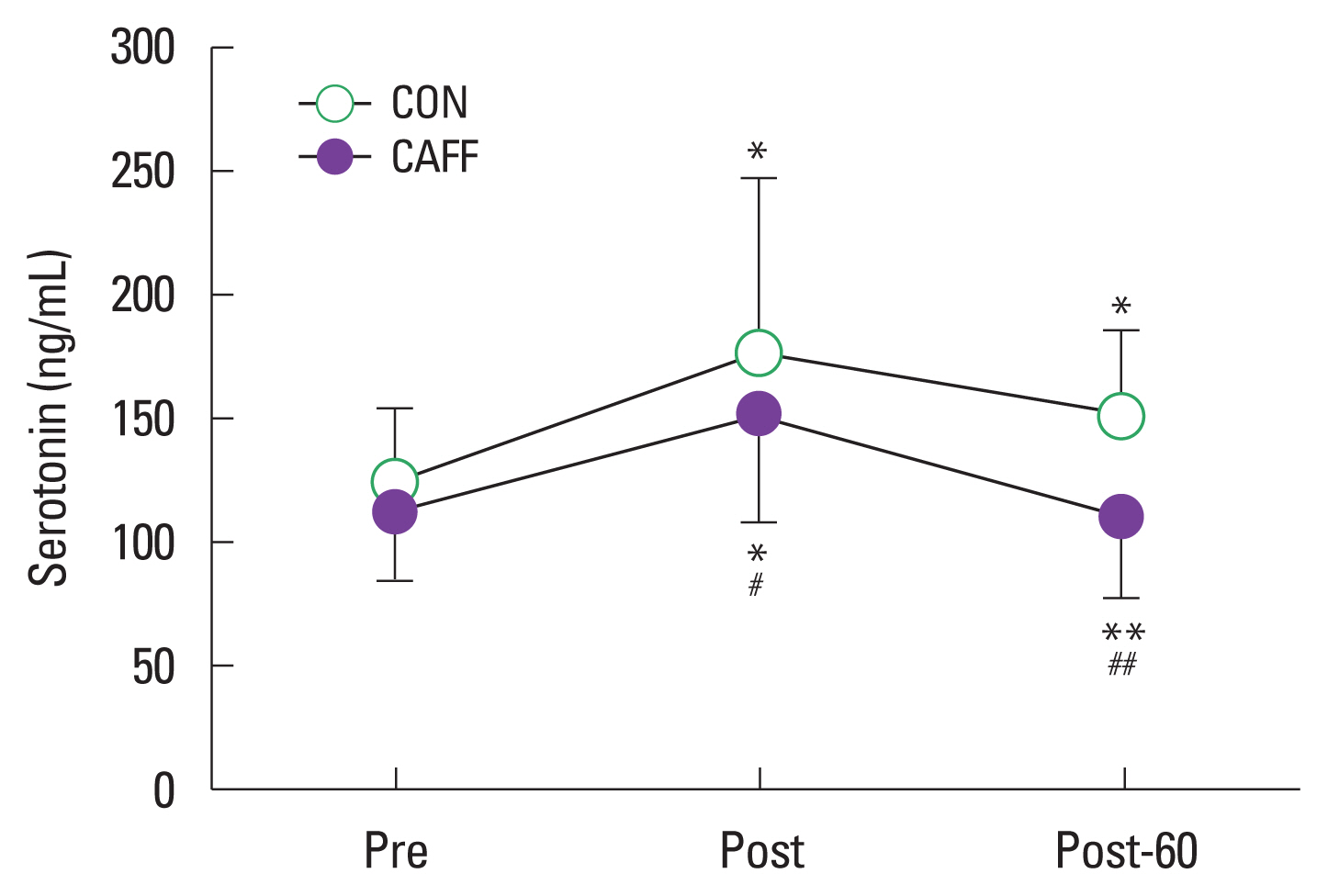
Serotonin
Serotonin increased significantly in both groups after passive heat loading for 30 min (Post, CAFF, P<0.05 vs. CON, P<0.05) and was significantly different in the CAFF and CON groups (150.70± 43.86 ng/mL vs. 174.90±71.27 ng/mL, P<0.05). Serotonin was significantly lower in CAFF than CON after 60 min of resting (Post-60, 109.10±33.67 ng/mL vs. 149.90±35.83 ng/mL, P< 0.01) (Fig. 1).
Dopamine
Dopamine increased significantly in both groups after passive heat loading for 30 min (Post, CAFF, P<0.001 vs. CON, P<0.05). Dopamine was significantly higher in CAFF than CON at the Post time point (16.00±6.70 pg/mL vs. 11.10±2.10 pg/mL, P< 0.001). Dopamine decreased significantly in only CAFF after 60 min of resting (P<0.01). However, there was no significant difference between CAFF and CON (Fig. 2).
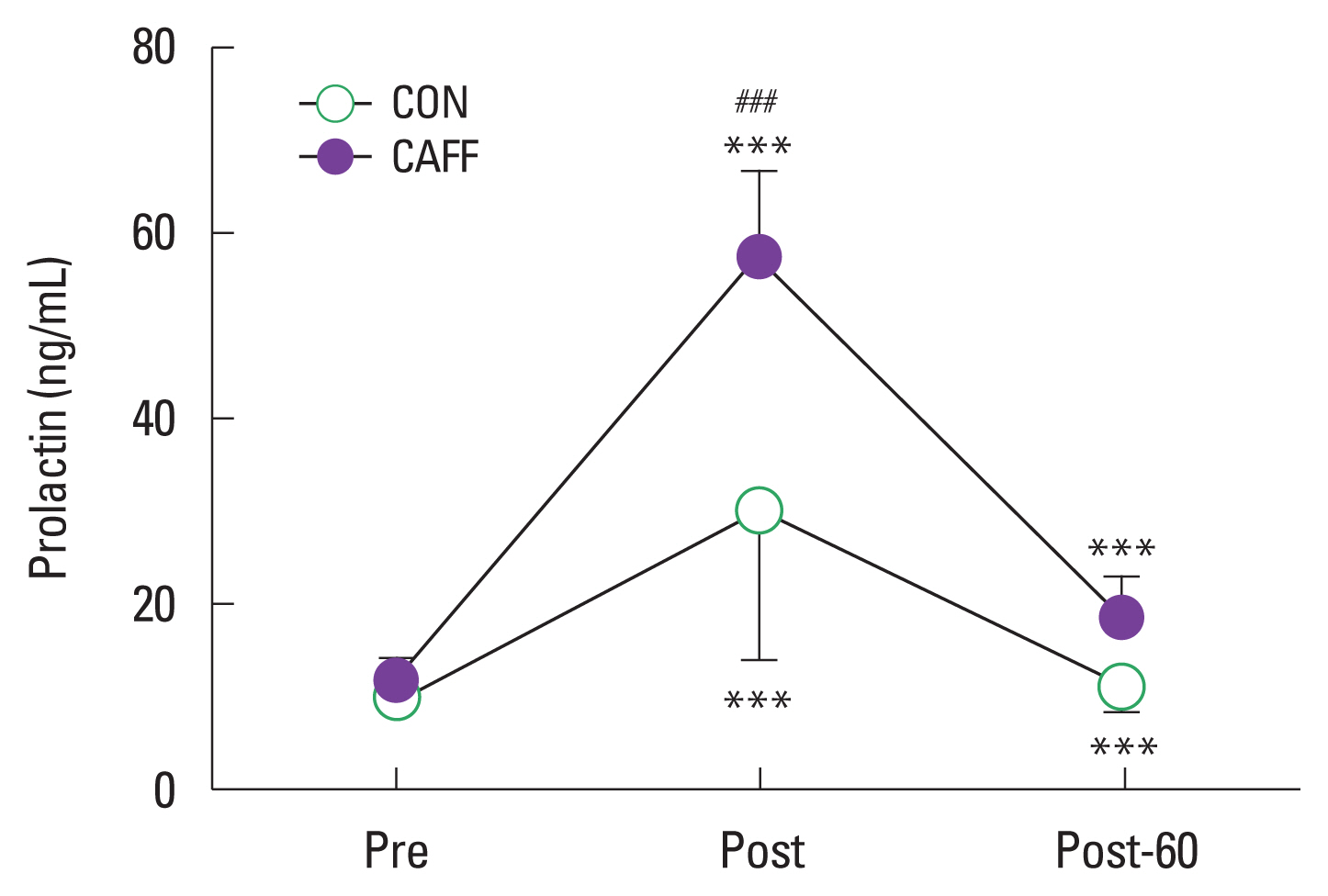
Prolactin
Prolactin increased significantly in both groups after passive heat loading for 30 min (Post, P<0.001). Post levels were significantly higher in CAFF than CON (57.47±9.14 ng/mL vs. 30.09± 16.00 ng/mL, P<0.001). After 60 min of resting, prolactin was significantly lower in both groups (P<0.001). However, there was no significant difference between CAFF and CON (Fig. 3).
DISCUSSION
This study evaluated a no caffeine group and found elevated serotonin, dopamine, and prolactin activities during passive heat loading, followed by a marked drop after 60 min of rest. However, a significantly lower level of serotonin and a higher level of dopamine and prolactin activity during passive heat loading were observed after caffeine ingestion.
It was previously reported that caffeine has the capability to reduce brain serotonin synthesis by inhibiting tryptophan hydroxylase, the rate-limiting enzyme for central serotonin biosynthesis (Lim et al., 2001), and/or to reduce brain serotonin/dopamine ratio by blocking adenosine α1 and α2 receptors within the CNS. Therefore, the effects of caffeine ingestion may be due to increased brain dopamine levels (Davis et al., 2003). Caffeine has been shown to cross the blood-brain barrier with the capability to serve as an antagonist of adenosine at the micromolar levels utilized in the previous study (Fredholm et al., 1999). Therefore, a significantly lower level of serotonin and a higher level of dopamine were observed after caffeine ingestion during passive heat loading in this study. These results would be attributed to increased central dopamine release due to caffeine antagonizing the inhibition of adenosine α1 and α2 receptors on dopamine, with a subsequent reduction in brain serotonin synthesis. Therefore, the results of this study suggested that central fatigue can decrease during passive heat loading and possibly occurs via alterations in the central serotonergic and dopaminergic activity from caffeine ingestion.
Peripheral secretion of prolactin has been suggested as a substitution measure of central fatigue during heat stress because blood levels tend to reflect the interaction between brain serotonergic and dopaminergic activity and prolactin has a strong connection with core temperature (Low et al., 2005). Therefore, prolactin is directly related to changes in body temperature. The increase in prolactin with core temperature in the previous study is consistent with the hypothesis associating central fatigue with decreased dopamine and increased serotonin (Wright et al., 2012).
It is difficult to account for why prolactin in the present experiment was at a higher level with caffeine ingestion than without despite the higher dopamine. The prolactin increased sharply in the caffeine ingestion group despite an increase in dopamine.
A previous study suggested that the inhibitory effects of dopamine on prolactin appeared to have been reduced during exertional heat stress (40°C, 30% relative humidity, wind speed <0.1 m/sec). Therefore, considering the passive heat loading temperature (42°C) and duration (30 min) in this study, the possibility of inhibitory effects of dopamine on prolactin by caffeine may be offset. This is similar to the previous study (Wright et al., 2012).
Although we did not present the results for the change in body temperature for the present study, in a previous study, we compared mean body temperature between the no caffeine and caffeine groups over the course of running for 30 min at 60% intensity of maximal oxygen consumption (experiments were conducted in a climate chamber at 24.0°C±0.5°C temperature, 40.0%±3.0% relative humidity, and <1-m/sec air velocity). The mean body temperature was significantly higher in the caffeine ingestion group than the no caffeine group at the end of the running period (Kim et al., 2011). However, the significance of the differences was negligible because mean body temperature was already at basal levels (before vs. after running; 36.26°C±0.14°C vs. 36.67°C±0.19°C). Therefore, caffeine can affect neurotransmitter release in the brain and the mechanism of caffeine on thermoregulatory might be due to neurotransmitter change in the preoptic and anterior hypothalamus (Zheng et al., 2014). And it would not cause excessive body temperature increase, leading to central fatigue.
Alteration in serotonin, dopamine, and prolactin under chronic passive heat loading or caffeine ingestion that produce heat or caffeine adaptation may follow a variant mechanism from changes under acute passive heat loading and caffeine. Therefore, change in central serotonergic and dopaminergic activities according to different stimulations are still issues to be clarified, including the effects of caffeine dose. In conclusion, 3-mg/kg caffeine ingestion can cause alterations in central serotonergic and dopaminergic activity during passive heat loading, which may contribute to reduced central fatigue and subsequently, to reduced fatigue. The prolactin response during passive heat loading was also significantly related to the caffeine ingestion in this study.