This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
Recent studies showed association between diseases and TLR6 polymorphisms. To investigate whether TLR6 polymorphisms are associated with the development of ischemic stroke, four single nucleotide polymorphisms (SNPs) of the TLR6 gene (rs1039559, rs3821985, rs3775073, and rs5743818) were analyzed in 120 patients with ischemic stroke (IS) and 278 control subjects. All ischemic stroke patients were classified into clinical subgroups according to NHISS and MBI. SNPStats was used to obtain odds ratios (ORs), 95% confidence intervals (CIs), and P values. Multiple logistic regression models (codominant1, codominant2, dominant, recessive, and log-additive) were performed to analyze the genetic data. Two SNPs (rs3821985 and rs3775073) of the TLR6 gene were associated with the NHISS in ischemic stroke patients (P< 0.05). Also, three SNPs (rs1039559, rs3821985, and rs3775073) showed association with MBI in ischemic stroke patients (P< 0.05). These results suggest that SNPs of TLR6 (rs1039559, rs3821985, and rs3775073) may be affect the disease characteristics of stroke, such as NIHSS and MBI.
The toll-like receptor (TLR) 6 has a role in the innate immunity, as receptor for pathogenic molecules (Kadowaki et al., 2001) and endogeneous damage-associated molecules (Piccinini et al., 2010). TLR6 is known as a receptor for mycobacterial infection (Zhang et al., 2013), and associated with recognization of bacterial lipopeptides and lipoteichoic acid (Irvine et al., 2013).
TLRs have receptor function for specific molecules (Kadowaki et al., 2001; Piccinini et al., 2010), and the overall expression of TLRs may activate the subsequent inflammatory response cascades (Kadowaki et al., 2001). TLRs that expressed on the cell membrane need to be dimerized to transduce signals into cytoplasmic pathways. Among them, TLR1 is the most expressed and may be dimerized with any other TLRs to transduce the recognized signals by other TLRs. For the TLR6, TLR2 is the major TLR dimers with it (Irvine et al., 2013).
Interestingly, there is a TLR10/TLR1/TLR6 locus on chromosome 4 (Mikacenic et al., 2013), and the locus forms a complex to genetically affect, or to be associated with, other immune response related genes. And it implies that TLR6 may have close genetic relationship to TLR1, the most expressed TLR which may affect the signal transduction of many TLRs. There was a report that the genetic variation in the TLR10/TLR1/TLR6 complex may determine the TLR1 and TLR2 mediated responses in the individuals (Mikacenic et al., 2013). And TLR1 and TLR2 is associated with activation of inflammatory responses in central nervous system, such as microglial cells, glial progenitors, neurons, astrocytes, as well as peripheral blood monocytes (Cassiani-Ingoni et al., 2006). In the Lyme disease, TLR1 and 2 may affect the human leukocyte antigen (HLA)-class II expressions and therefore, differential symptoms, such as whether central nervous system manifestation develops in individuals, may be determined. Moreover, TNF-alpha mediated inflammation may contribute to the cerebral aneurysms, which is another cerebral artery disease and related with severe inflammatory responses, and TLR-mediated innate immunity was related with higher TNF-alpha expression (Jayaraman et al., 2008). Such evidences suggest that TLRs may affect many vascular diseases, such as stroke, however TLR6 was mostly studied in the pulmonary diseases, such as bronchiolitis (Nuolivirta et al., 2013), Legionnaires’ disease (Misch et al., 2013), and bronchial effect of ureaplasma (Winters et al., 2013), but no previous study was reported whether the TLR6 polymorphisms may affect the development or clinical features of stroke.
Therefore, we investigated associations between the single nucleotide polymorphism (SNP) of TLR6 and development or clinical features of ischemic stroke.
MATERIALS AND METHODS
Study subjects
Stroke patients were recruited from Kyung Hee Medical Center. Table 1 shows the clinical characteristics study participants. The controls were 278 aged male and female (148 male and 130 female, mean age 63.6 yr) without any severe diseases. And the stroke patients were 120 aged male and female (67 male and 53 female, mean age 65.8 yr), and their clinical characteristics such as National Institutes of Health Stroke Scale (NIHSS) and modified barthel index (MBI) were analyzed. Ethical approval of this study was obtained from the ethics review committee of Medical Research Institute, School of Medicine, Kyung Hee University, Seoul, Korea.
SNP selection and genotyping
We selected four SNPs in TLR6: rs1039559 (intron), rs3821985 (Thr361Thr), rs3775073 (Lys421Lys), and rs5743818 (Ala644A-la). They were searched from the SNP database in NCBI (http://www.ncbi.nlm.nih.gov/SNP), and reviewed to select the region in exons and near exons. Peripheral blood of all patients were sampled in EDTA blood tube, stored in −20°C freezer before the extraction of genomic DNA. Genomic DNA was extracted with QIAamp® DNA extraction kit (QIAGEN, Valencia, CA, USA). Polymerase chain reactions (PCRs) of the SNPs were performed using the following primers in the Table 2 to amplify target sequences including the SNPs. The primers used in this study were in a previous study of TLRs (Kim et al., 2013). Direct sequencing, sequence sorting, and analyzing were done with ABI PRISM 3730XL analyzer (PE Applied Biosystems, Foster City, CA, USA) and with SeqManII software (DNASTAR, Madison, WI, USA).
Statistical analysis
SNPStats (http://bioinfo.iconcologia.net/index.php?module=Snpstats) and SPSS 20.0 (SPSS Inc., Chicago, IL, USA) were used to obtain odds ratios (ORs), 95% confidence intervals (CIs), and P values. Multiple logistic regression models were applied in the analysis of genotypes. Each component of the model was: codominant1 (major allele homozygotes vs heterozygotes), codominant2 (major allele homozygotes vs minor allele homozygotes), dominant (major allele homozygotes vs heterozygotes+minor allele homozygotes), recessive (major allele homozygotes+heterozygotes vs. minor allele homozygotes), and log-additive (major allele homozygotes vs. heterozygotes vs. minor allele homozygotes) (Kim et al., 2013). Age and gender were included as covariates and adjusted to obtain statistical significance. The linkage disequilibrium (LD) block and haplotypes between pairs of SNPs were tested with Haploview 4.2 software (Daly Lab, Cambridge, MA, USA). In the statistical analysis, P<0.05 was considered significant. Hardy-Weinberg equilibrium of each SNP was tested in SNPstats.
RESULTS
The frequencies of genotype and allele of TLR6 in control and patients with ischemic stroke were displayed in the Table 3, however, there was no significantly associated with development of ischemic stroke (P>0.05). All of the four tested SNPs were in Hardy-Weinberg equilibrium (data not shown).
The genotype and allele frequencies of SNPs of TLR6 gene in ischemic stroke patients with NIHSS score (<6) and ICH patients with NIHSS score (≥6) show in the Table 4. For the two of synonymous SNPs rs3821985 (Thr361Thr) and rs3775073 (Lys421-Lys), the genotype distributions were associated with NIHSS (rs-3821985 and rs3775073, OR=0.40, 95% CI=0.18–0.90, P= 0.026 in codominant1, OR=0.40, 95% CI=0.19–0.88, P=0.02 in dominant, and OR=0.54, 95% CI=0.30–0.99, P=0.043 in log-additive).
The allele frequencies of rs3821985 and rs3775073 were also associated with NIHSS (OR=0.57, 95% CI=0.32–0.99, P=0.048), in both SNPs. However, an intronic SNP rs1039559 and a synonymous SNP rs5743818 (Ala644Ala) was not associated with NIHSS in this study.
The genotype and allele frequencies of SNPs of TLR6 gene in ischemic stroke patients with MBI score (<60) and ischemic stroke patients with MBI score (≥60) were shown in the Table 5. An intronic SNP rs1039559 was associated with MBI score of stroke patients (OR=0.30, 95% CI=0.10–0.90, P=0.032 in codominant, OR=0.27, 95% CI=0.09–0.81, P=0.014 in dominant, and OR=0.28, 95% CI=0.10–0.80, P=0.0096 in log-additive). And the two of synonymous SNPs rs3821985 (Thr361Thr) and rs3775073 (Lys421Lys) also were associated with MBI in codominant1 (OR=3.36, 95% CI=1.07–10.60, P=0.038) model.
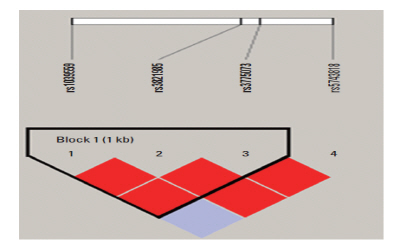
There was a LD block between only three (rs1039559, rs3821985, and rs3775073) of the four tested SNPs (Fig. 1), and the LD block was strong between rs3821985 and rs3775073 (D’=1.0, r-squared=1.0). Therefore, haplotype analysis was performed; however, no haplotype was significantly associated with the development of ischemic stroke (P>0.05).
DISCUSSION
TLR6 is located on the chromosome 4, forming a locus of TLR10/TLR1/TLR6 complex. The locus is associated with many relatively common diseases, such as papillary thyroid carcinoma (Kim et al., 2013), sarcoidosis (Veltkamp et al., 2012), Crohn’s disease (Abad et al., 2011), and various cancer risks (Kutikhin, 2011).
Function of TLR6 may be disturbed by genetic defects (Frazao et al., 2013). Changes in the genome of individual may contribute to the altered innate immune responses, and auch influence may involve inflammatory cytokines and chemokines, such as IL-1β, IL-6, IL-8, IL-12, and TNF-α (Frazão et al., 2013).
Inflammatory mediators are important in stroke, because vascular inflammations may lead to the development of stroke (Chistiakov et al., 2013), and moreover, responses of such genes are related to the pathologic process after the hemorrhage occurs (Altay et al., 2013; Ostergaard et al., 2013; Zhou et al., 2013). NLRP3 inflammasome is a major component of the innate immune system, which affects the neutrophil function and edematous tissue responses (Ma et al., 2013). The action of inflammasome involves the caspase-1, IL-1b, moreover, it is associated with activation of TLRs (Fernandes-Alnemri et al., 2013). Tissue hypoxia caused by ischemic changes in stroke (Ostergaard et al., 2013) may lead to ischemia-induced inflammatory responses, which was well-investigated in cardiovascular diseases (Guo et al., 2013) and cerebrovascular diseases (Taylor et al., 2013). In cerebrovascular disease models, microglial activation is an important phenomenon in ischemic inflammatory responses, and TLR6 was reported to be associated with microglial responses. TLR6 with TLR4 forms a dimer and it may promote inflammations by CD36 ligand, which is also related to oxidative injury and hemorrhagic disorders (Olson et al., 2004).
In our result, two exonic SNPs rs3821985 and rs3775073 were associated with the clinical features (NIHSS and MBI) of stroke, and an intronic SNP rs1039559 was associated with MBI. Interestingly, only intronic SNP rs1039559 was previously reported to be associated with higher total immunoglobulin (Ig) E levels, especially in the children aged 1–2 yr (Reijmerink et al., 2011). There have been some previous studies of Ser249Pro, a famous SNP in TLR6, however, reported only association with asthmatic disorders (Hoffjan et al., 2005). Therefore, it may be the first study to report the association between TLR6 and stroke.
In conclusion, we suggest that SNPs of TLR6 (rs1039559, rs-3821985, and rs3775073) may affect the disease characteristics of ischemic stroke, such as NIHSS and MBI. Additionally, MBI is a score related to the functional assessment of rehabilitation, our result may contribute to the genetic study of rehabilitation fields. Our study has some limitations, such as only small population of Korean were analyzed. Therefore, more subjects will be needed to confirm our results in further study.
Notes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.
REFERENCES
Abad C, González-Escribano MF, Diaz-Gallo LM, Lucena-Soto JM, Márquez JL, Leo E, Crivell C, Gómez-García M, Martín J, Núñez-Roldán A, García-Lozano JR. Association of Toll-like receptor 10 and susceptibility to Crohn’s disease independent of NOD2. Genes Immun. 2011;12:635–642.
Altay O, Suzuki H, Hasegawa Y, Ostrowski RP, Tang J, Zhang JH. Isoflurane on brain inflammation. Neurobiol Dis. 2013;62C:365–371.
Cassiani-Ingoni R, Cabral ES, Lunemann JD, Garza Z, Magnus T, Gelderblom H, Munson PJ, Marques A, Martin R. Borrelia burgdorferi induces TLR1 and TLR2 in human microglia and peripheral blood monocytes but differentially regulates HLA-class II expression. J Neuropathol Exp Neurol. 2006;65:540–548.
Chistiakov DA, Sobenin IA, Orekhov AN. Vascular extracellular matrix in atherosclerosis. Cardiol Rev. 2013;21:270–288.
Fernandes-Alnemri T, Kang S, Anderson C, Sagara J, Fitzgerald KA, Alnemri ES. Cutting edge: TLR signaling licenses IRAK1 for rapid activation of the NLRP3 inflammasome. J Immunol. 2013;191:3995–3999.
Frazão JB, Errante PR, Condino-Neto A. Toll-like receptors’ pathway disturbances are associated with increased susceptibility to infections in humans. Arch Immunol Ther Exp (Warsz). 2013;61:427–443.
Guo J, Wang SB, Yuan TY, Wu YJ, Yan Y, Li L, Xu XN, Gong LL, Qin HL, Fang LH, Du GH. Coptisine protects rat heart against myocardial ischemia/reperfusion injury by suppressing myocardial apoptosis and inflammation. Atherosclerosis. 2013;231:384–391.
Hoffjan S, Stemmler S, Parwez Q, Petrasch-Parwez E, Arinir U, Rohde G, Reinitz-Rademacher K, Schultze-Werninghaus G, Bufe A, Epplen JT. Evaluation of the toll-like receptor 6 Ser249Pro polymorphism in patients with asthma, atopic dermatitis and chronic obstructive pulmonary disease. BMC Med Genet. 2005;6:34
Irvine KL, Hopkins LJ, Gangloff M, Bryant CE. The molecular basis for recognition of bacterial ligands at equine TLR2, TLR1 and TLR6. Vet Res. 2013;44:50
Jayaraman T, Paget A, Shin YS, Li X, Mayer J, Chaudhry H, Niimi Y, Silane M, Berenstein A. TNF-alpha-mediated inflammation in cerebral aneurysms: a potential link to growth and rupture. Vasc Health Risk Manag. 2008;4:805–817.
Kadowaki N, Ho S, Antonenko S, Malefyt RW, Kastelein RA, Bazan F, Liu YJ. Subsets of human dendritic cell precursors express different toll-like receptors and respond to different microbial antigens. J Exp Med. 2001;194:863–869.
Kim SK, Park HJ, Hong IK, Chung JH, Eun YG. A missense polymorphism (rs11466653, Met326Thr) of toll-like receptor 10 (TLR10) is associated with tumor size of papillary thyroid carcinoma in the Korean population. Endocrine. 2013;43:161–169.
Kutikhin AG. Association of polymorphisms in TLR genes and in genes of the Toll-like receptor signaling pathway with cancer risk. Hum Immunol. 2011;72:1095–1116.
Ma Q, Chen S, Hu Q, Feng H, Zhang JH, Tang J. NLRP3 inflammasome contributes to inflammation after intracerebral hemorrhage. Ann Neurol. 2013;10.1002/ana.24070.
Mikacenic C, Reiner AP, Holden TD, Nickerson DA, Wurfel MM. Variation in the TLR10/TLR1/TLR6 locus is the major genetic determinant of interindividual difference in TLR1/2-mediated responses. Genes Immun. 2013;14:52–57.
Misch EA, Verbon A, Prins JM, Skerrett SJ, Hawn TR. A TLR6 polymorphism is associated with increased risk of Legionnaires’ disease. Genes Immun. 2013;14:420–426.
Nuolivirta K, Vuononvirta J, Peltola V, Koponen P, Helminen M, He Q, Korppi M. Toll-like receptor-2 subfamily genotypes are not associated with severity of bronchiolitis or post-bronchiolitis wheezing in infants. Acta Paediatr. 2013;10.1111/apa.12425.
Olson JK, Miller SD. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs. J Immunol. 2004;173:3916–3924.
Ostergaard L, Aamand R, Karabegovic S, Tietze A, Blicher JU, Mikkelsen IK, Iversen NK, Secher N, Engedal TS, Anzabi M, Jimenez EG, Cai C, Koch KU, Næss-Schmidt ET, Obel A, Juul N, Rasmussen M, Sørensen JC. The role of the microcirculation in delayed cerebral ischemia and chronic degenerative changes after subarachnoid hemorrhage. J Cereb Blood Flow Metab. 2013;33:1825–1837.
Piccinini AM, Midwood KS. DAMPening inflammation by modulating TLR signalling. Mediators Inflamm. 2010;2010:672395
Reijmerink NE, Kerkhof M, Bottema RW, Gerritsen J, Stelma FF, Thijs C, van Schayck CP, Smit HA, Brunekreef B, Postma DS, Koppelman GH. Toll-like receptors and microbial exposure: gene-gene and gene-environment interaction in the development of atopy. Eur Respir J. 2011;38:833–840.
Taylor RA, Sansing LH. Microglial responses after ischemic stroke and intracerebral hemorrhage. Clin Dev Immunol. 2013;2013:746068
Veltkamp M, van Moorsel CH, Rijkers GT, Ruven HJ, Grutters JC. Genetic variation in the Toll-like receptor gene cluster (TLR10-TLR1-TLR6) influences disease course in sarcoidosis. Tissue Antigens. 2012;79:25–32.
Winters AH, Levan TD, Vogel SN, Chesko KL, Pollin TI, Viscardi RM. Single nucleotide polymorphism in toll-like receptor 6 is associated with a decreased risk for ureaplasma respiratory tract colonization and bronchopulmonary dysplasia in preterm infants. Pediatr Infect Dis J. 2013;32:898–904.
Zhang Y, Jiang T, Yang X, Xue Y, Wang C, Liu J, Zhang X, Chen Z, Zhao M, Li JC. Toll-like receptor -1, -2, and -6 polymorphisms and pulmonary tuberculosis susceptibility: a systematic review and meta-analysis. PLoS One. 2013;8:e63357
Zhou QB, Jin YL, Jia Q, Zhang Y, Li LY, Liu P, Liu YT. Baicalin attenuates brain edema in a rat model of intracerebral hemorrhage. Inflammation. 2013;10.1007/s10753-013-9717-9.
Fig. 1.
Linkage disequilibrium block consists of rs1039559, rs3821985, and rs3775073.
Table 1.
Clinical characteristics in IS and control subjects
IS
Control
Total number
120
278
Male/female (n)
67/53
148/130
Age (mean age ± SD)
65.8± 12.1
63.6± 9.2
NIHSS (score)
< 6
56
≥ 6
56
MBI (score)
< 60
70
≥ 60
25
Stroke patients with inappropriate clinical data were excluded. IS, ischemic stroke; SD, standard deviation; NIHSS, National Institutes of Health Stroke Survey; MBI, Modified Barthel Index.
Table 2.
Primer sequences for each SNP
SNPs
Sense (5′-3′)
Anti-sense (5′-3′)
Size (bp)
rs1039559
GTGGTTGTGTGTTTTGACCTGT
TGTTGGATCACTTTCTCAATGC
452
rs3821985
TTGAAAGCATTCGTGAAGAAGA
TGATCCTGGGAGGTAAACATCT
492
rs3775073
TACCTTGATCCTGGGAGGTAAA
TTGAAAGCATTCGTGAAGAAGA
497
rs5743818
AGCCCACTAAAGGACTTTCACA
GGGAGACAAAACAAAGATGGAC
405
SNP, single nucleotide polymorphism.
Table 3.
Frequencies of genotype and allele of TLR6 in control and patients with IS
SNP
Genotype/allele
Control
IS
Models
OR (95% CI)
P
n (%)
n (%)
rs1039559
T/T
141 (50.7)
64 (53.3)
Codominant1
0.91 (0.58–1.43)
0.69
Intron
T/C
118 (42.5)
49 (40.8)
Codominant2
0.78 (0.31–1.97)
0.60
C/C
19 (6.8)
7 (5.8)
Dominant
0.90 (0.58–1.38)
0.61
Recessive
0.82 (0.33–2.01)
0.65
Log-additive
0.90 (0.63–1.28)
0.56
T
400 (71.9)
177 (73.8)
1
C
156 (28.1)
63 (26.2)
0.91 (0.65–1.29)
0.60
rs3821985
C/C
146 (52.5)
54 (45.0)
Codominant1
1.38 (0.88–2.18)
0.16
Thr361Thr
C/G
105 (37.8)
54 (45.0)
Codominant2
1.20 (0.56–2.55)
0.64
G/G
27 (9.7)
12 (10.0)
Dominant
1.35 (0.87–2.07)
0.18
Recessive
1.03 (0.50–2.13)
0.93
Log-additive
1.19 (0.86–1.64)
0.29
C
397 (71.4)
162 (67.5)
1
G
159 (28.6)
78 (32.5)
1.20 (0.87–1.67)
0.27
rs3775073
A/A
146 (52.5)
54 (45.0)
Codominant1
1.38 (0.88–2.18)
0.16
Lys421Lys
A/G
105 (37.8)
54 (45.0)
Codominant2
1.20 (0.56–2.55)
0.64
G/G
27 (9.7)
12 (10.0)
Dominant
1.35 (0.87–2.07)
0.18
Recessive
1.03 (0.50–2.13)
0.93
Log-additive
1.19 (0.86–1.64)
0.29
A
397
162
1
G
159
78
1.20 (0.87–1.67)
0.27
rs5743818
T/T
265 (95.3)
117 (97.5)
Codominant1
0.57 (0.16–2.06)
0.37
Ala644Ala
T/G
13 (4.7)
3 (2.5)
T
543 (97.7)
237 (98.8)
1
G
13 (2.3)
3 (1.2)
0.53 (0.15–1.87)
0.42c
The P values were calculated from logistic regression analysis adjusting sex and age. The Pc was calculated using Fisher’s exact test. IS, ischemic stroke; SNP, singe nucleotide polymorphism; OR, odds ratio; CI, confidence interval.
Table 4.
Genotype and allele frequencies of SNPs of TLR6 gene in IS patients with NIHSS score (< 6) and ICH patients with NIHSS score (≥ 6)
SNP
Genotype/allele
NHISS (< 6)
NHISS (≥ 6)
Models
OR (95% CI)
P
n (%)
n (%)
rs1039559
T/T
33 (58.9)
25 (44.6)
Codominant1
1.91 (0.87–4.18)
0.11
Intron
T/C
19 (33.9)
28 (50.0)
Codominant2
0.94 (0.19–4.67)
0.94
C/C
4 (7.1)
3 (5.4)
Dominant
1.74 (0.82–3.71)
0.15
Recessive
0.70 (0.15–3.32)
0.65
Log-additive
1.37 (0.74–2.55)
0.31
T
85 (75.9)
78 (69.6)
1
C
27 (24.1)
34 (30.4)
1.37 (0.76–2.48)
0.29
rs3821985
C/C
19 (33.9)
31 (55.4)
Codominant1
0.40 (0.18–0.90)
0.026
Thr361Thr
C/G
30 (53.6)
20 (35.7)
Codominant2
0.43 (0.11–1.64)
0.22
G/G
7 (12.5)
5 (8.9)
Dominant
0.40 (0.19––0.88)
0.02
Recessive
0.71 (0.20–2.47)
0.58
Log-additive
0.54 (0.30–0.99)
0.043
C
68 (60.7)
82 (73.2)
1
G
44 (39.3)
30 (26.8)
0.57 (0.32–0.99)
0.048
rs3775073
A/A
19 (33.9)
31 (55.4)
Codominant1
0.40 (0.18–0.90)
0.026
Lys421Lys
A/G
30 (53.6)
20 (35.7)
Codominant2
0.43 (0.11–1.64)
0.22
G/G
7 (12.5)
5 (8.9)
Dominant
0.40 (0.19–0.88)
0.02
Recessive
0.71 (0.20–2.47)
0.58
Log-additive
0.54 (0.30–0.99)
0.043
A
68 (60.7)
82 (73.2)
1
G
44 (39.3)
30 (26.8)
0.57 (0.32–0.99)
0.048
rs5743818
T/T
53 (94.6)
56 (100.0)
Codominant1
0.00 (0.00-NA)
NA
Ala644Ala
T/G
3 (5.4)
0 (0.0)
T
109 (97.3)
112 (100.0)
G
3 (2.7)
0 (0.0)
0.25c
The P values were calculated from logistic regression analysis adjusting sex and age. The Pc was calculated using Fisher’s exact test. Bold numbers indicate significant associations. IS, ischemic stroke; SNP, singe nucleotide polymorphism; NIHSS, National Institutes of Health Stroke Survey; OR, odds ratio; CI, confidence interval; NA, not applicable.
Table 5.
Genotype and allele frequencies of SNPs of TLR6 gene in IS patients with MBI score (< 60) and IS patients with MBI score (≥ 60)
SNP
Genotype/allele
MBI (< 60)
MBI (≥ 60)
Models
OR (95% CI)
P
n (%)
n (%)
rs1039559
T/T
33 (47.1)
19 (76.0)
Codominant1
0.30 (0.10–0.90)
0.032
Intron
T/C
33 (47.1)
6 (24.0)
Codominant2
0.00 (0.00-NA)
NA
C/C
4 (5.7)
0 (0.0)
Dominant
0.27 (0.09–0.81)
0.014
Recessive
0.00 (0.00-NA)
NA
Log-additive
0.28 (0.10–0.80)
0.0096
T
99 (70.7)
44 (88.0)
1
C
41 (29.3)
6 (12.0)
0.33 (0.13–0.83)
0.019
rs3821985
C/C
32 (45.7)
7 (28.0)
Codominant1
3.36 (1.07–10.60)
0.038
Thr361Thr
C/G
30 (42.9)
16 (64.0)
Codominant2
0.57 (0.09–3.54)
0.55
G/G
8 (11.4)
2 (8.0)
Dominant
2.27 (0.79–6.52)
0.12
Recessive
0.31 (0.06–1.68)
0.14
Log-additive
1.16 (0.57–2.37)
0.68
C
94 (67.1)
30 (60.0)
1
G
46 (32.9)
20 (40.0)
1.36 (0.70–2.65)
0.36
rs3775073
A/A
32 (45.7)
7 (28.0)
Codominant1
3.36 (1.07–10.60)
0.038
Lys421Lys
A/G
30 (42.9)
16 (64.0)
Codominant2
0.57 (0.09–3.54)
0.55
G/G
8 (11.4)
2 (8.0)
Dominant
2.27 (0.79–6.52)
0.12
Recessive
0.31 (0.06–1.68)
0.14
Log-additive
1.16 (0.57–2.37)
0.68
A
94 (67.1)
30 (60.0)
1
G
46 (32.9)
20 (40.0)
1.36 (0.70–2.65)
0.36
rs5743818
T/T
67 (95.7)
25 (100.0)
Codominant1
0.00 (0.00-NA)
NA
Ala644Ala
T/G
3 (4.3)
0 (0.0)
T
137 (97.9)
50 (100.0)
G
3 (2.1)
0 (0.0)
0.57c
The P values were calculated from logistic regression analysis. The Pc was calculated using Fisher’s exact test. Bold numbers indicate significant associations. IS, ischemic stroke; SNP, singe nucleotide polymorphism; MBI, Modified Barthel Index; OR, odds ratio; CI, confidence interval; NA, not applicable.