INTRODUCTION
Hypoxic ischemia brain injury is a common cause of morbidity and mortality in neonates (Demers et al., 2005). Hypoxic ischemia brain injury in neonates results in severe encephalopathy, seizures, and cerebral palsy in survived infants (Brucknerová et al., 2006; Chalmers, 2005; Obenaus and Ashwal, 2008).
The nigro-striatal dopaminergic pathway extends from the substantia nigra to the dorsal striatum via the median forebrain bundle (Bergquist et al., 2003). The substantia nigra plays important roles in eye movement, motor planning, reward seeking, learning, and addiction. The most functions of substantia nigra are mediated through the striatum, because the nigral dopaminergic input to the striatum occurs via the nigrostriatal pathway (Nicola et al., 2000). Dopamine neurons in the substantia nigra are vulnerable to the hypoxic-ischemic conditions and undergo degeneration (Singh et al., 2007). Striatum is also one of the most sensitive brain areas to the hypoxic ischemia injury (González-Hernández and Rodríguez, 2000).
Dopamine is a prevalent catecholaminergic neurotransmitter in the brain, and dopaminergic neurons are mainly localized in the substantia nigra pars compacta (Nelson et al., 1996). Dopamine plays a significant role in memory processes, especially through the interconnection of the striatum and the prefrontal cortex (Jay, 2003). In acute brain slices, these dopamine neurons undergo hyperpolarization during hypoxia and hypoglycemia, which results in silencing of the neurons (Singh et al., 2007). Following striatal hypoxic ischemica injury, there was a reduction in the number of substantia nigra dopaminergic neurons. These reductions occurred in the absence of apparent direct hypoxic ischemia injury to the substantia nigra (Burke et al., 1992).
As dopamine and dopaminergic neurocircuitries are extremely sensitive to hypoxia and ischemia at birth (Klawitter et al., 2007), perinatal or postnatal hypoxic ischemia brain injury is associated with dopaminergic neuronal loss, and resulted in cognitive and motor impairments (Andreeva et al., 2001; Meng et al., 2006). Nigro-striatal dopamine disruption impairs the learning of complex motor tasks (Russell, 2003; Sullvian and Brake, 2003).
Tyrosine hydroxylase (TH) is a rate-limiting enzyme for the bio-synthesis of dopamine (Haavik and Toska, 1998). Oo et al. (1995) and Burke et al. (1992) indicated that postnatal 7 days neonatal rats exposed to the hypoxic ischemia injury induced by unilateral carotid ligation and followed by exposure to 8% O2 conditions for 4 h showed decrease in the number of TH-positive neurons in the substantia nigra.
Exercise improves spatial learning ability, memory capability, cognitive function, neurogenesis, and expressions of neurotrophic factors (O’Callaghan et al., 2007; Seo et al., 2013; Vasuta et al., 2007; Yasuhara et al., 2007). However, the effect of treadmill exercise on dopamine expression in the nigro-stratal dopaminergic pathway following hypoxic ischemia brain injury is currently unknown. In the present study, we investigated the effects of tread-mill exercise during adolescence on spatial learning ability in relation with nigro-striatal dopamine level following hypoxic ischemia brain injury in neonatal rats.
MATERIALS AND METHODS
Animals and treatments
Seven-day-old Sprague-Dawley rats (10–15 g weight) were used in this experiment. The experimental procedures were performed in accordance with the animal care guidelines of National Institutes of Health (NIH) and Korean Academy of Medical Sciences. The animals were housed under the controlled temperature (20± 2°C) and lighting conditions (07:00 to 19:00), with food and water made available ad libitum throughout the experiment. The animals were divided into the four groups (n=10 in each group): the sham-operation group, the sham-operation and exercise group, the hypoxic ischemia-induction group, and the hypoxic ischemia-induction and exercise group.
Induction of hypoxic ischemia brain injury
We used the hypoxic ischemia injury model introduced by Rice-Vannucci model (Rice et al., 1981). On postnatal 7 day, the neonatal rats were anesthetized with diethyl ether and then the midline of the neck was incised at the longitudinal plane. The left common carotid artery was exposed, dissected from the vagus nerve, and doubly ligated with 4.0 suture silk to ensure cessation of blood flow. Two hours after surgery, the neonatal rats were then exposed to hypoxia conditions (8% oxygen, 92% nitrogen) for one hour by placing them in an airtight plastic box which was provided a 37°C stable thermal environment. For the neonatal rats in the sham-operation group and in the sham-operation and exercise group, surgical incision was conducted without ligation, and exposure to hypoxic condition was deducted. After exposure to hypoxia condition, all neonatal rats were left undisturbed together with their respective mothers for 21 days.
Treadmill exercise protocol
The rat pups in the exercise groups were forced to run on a motorized treadmill for 30 min once a day for 12 weeks, starting 22 days after induction of hypoxic ischemia brain injury. The exercise load used in the first 2 weeks consisted of running at 2 meters/min, at 0 degree of inclination. Then, the speed of treadmill running was increased 2 meters/min each two weeks until 12 weeks, at 0 degree of inclination. The rat pups in the non-exercise groups were left in treadmill without running for the same period as the exercise groups.
Morris water maze test
In order to evaluate the spatial learning ability in rat pups, the latency in the Morris water maze test was determined, as the previously described method (Li et al., 2007). The water maze was a circular pool (diameter=140 cm, height=45 cm) constructed of fiberglass. The water was maintained at a temperature of 22±2°C. During testing in the water maze, a platform (diameter=15 cm, height=35 cm) was located 2 cm below the water surface in one of four locations in the pool. Clearly visible cues outside the basin were provided for orientation. The test consisted of three acquisition phases and two probe trials. In the acquisition phase, the rat pups had one daily session of four trials for 3 consecutive days to find the platform (diameter=15 cm, height=35 cm) submerged 2 cm under the water surface. For each trial, the rat pup was placed in the water, facing the wall of the tank, in one of the four start locations. The rat pup was allowed to search for the platform for 60 sec. If the rat pup found the platform, remaining on the platform for 10 sec was permitted. If the rat pup did not find the platform within 60 sec, the rat pup was guided and allowed to remain on the platform for 10 sec. The interval between trials was 20 sec. After each training session, the rat pups were dried with a towel and returned to their cages. To assess spatial learning ability, the animals were subjected to the 60 sec probe trial following the last training session, and then the platform was removed from the pool. The occupancy times in the quadrant platform were recorded automatically by a video tracking system (SMART; Pan-Lab, Barcelona, Spain).
Tissue preparation
The rat pups were sacrificed after the completion of last tread-mill exercise on the 12 weeks of the experiment. The animals were anesthetized using Zoletil 50® (10 mg/kg, i.p.; Vibac Laboratories, Carros, France). The rat pups were transcardially perfused with 50 mM phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde in 100 mM phosphate buffer (PB, pH 7.4). The brains were dissected and postfixed in the same fixative overnight and transferred into a 30% sucrose solution for cryoprotection. Coronal sections of 40 μm thickness in each section of brain was made with a freezing microtome (Leica, Nussloch, Germany).
Immunohistochemistry for TH
Immunohistochemistry was conducted to evaluate the TH expressions in the substantia nigra and striatum, according to the previously described method (Sung et al., 2012). An average of 10 sections within the substantia nigra spanning from Bregma −5.2 mm to −5.6 mm and within striatum spanning from Bregma −0.7 mm were obtained from each brain. The staining was carried out using free-gloating sections. The sections were incubated in PBS for 5 min and then washed three times in the same buffer. The sections were then incubated in 1% hydrogen peroxide (H2O2) for 30 min to block the endogenous peroxidase activity. After washing in PBS, the sections were incubated in blocking serum (10% normal horse serum and 0.1% Triton X-100 in PBS) for 2 h. Next, the sections were incubated overnight with the anti-TH monoclonal antibody solution (1:1,000; BD Pharmingen, San Diego, CA, USA). The sections were then incubated for 2 h with the biotinylated anti-mouse secondary antibody (1:300; Vector Laboratories, Burlingame, CA, USA). Next, the sections were incubated with avidin-biotin-peroxidase complex (Vector Laboratories) for 1 h at room temperature. For staining, the sections were incubated in a solution consisting of 0.02% 3,3′-diaminobenzidine tetrahydrochloride (DAB) and 0.03% H2O2 in 50 mM Tris-hydrochloride (HCl) (pH 7.6) for approximately 3 min, after which they were then washed with PBS and mounted onto gelatin-coated slides. The number of TH-positive cells in the substantia nigra was counted in each section using a microscope (Olympus, Tokyo, Japan), and the optical density of TH-fiber in the striatum was measured in images of the dorsolateral striatum Image-Pro Plus (Media Cybernetics, Bethesda, MD, USA).
Statistical analysis
All data were analyzed using the statistical software SPSS (version 16.0). The data were expressed as the mean±standard error of the mean (SEM). For the comparison among the groups, oneway ANOVA and Duncan’s post-hoc test were performed with P<0.05 as an indication of statistical significance.
RESULTS
Effect of treadmill exercise on spatial learning ability
Spatial learning ability was measured using the Morris-water maze test (Fig. 1). The time spent in the probe quadrant was 14.90 ±2.40 sec in the sham-operation group, 21.70±2.95 sec in the sham-operation and exercise group, 9.46±2.82 sec in the hypoxic ischemia-induction group, and 17.26±1.53 sec in the hypoxic ischemia-induction and exercise group. The present results showed that spatial learning ability was disturbed by induction of hypoxic ischemia injury and that treadmill exercise alleviated hypoxic ischemia-induced spatial learning impairment.
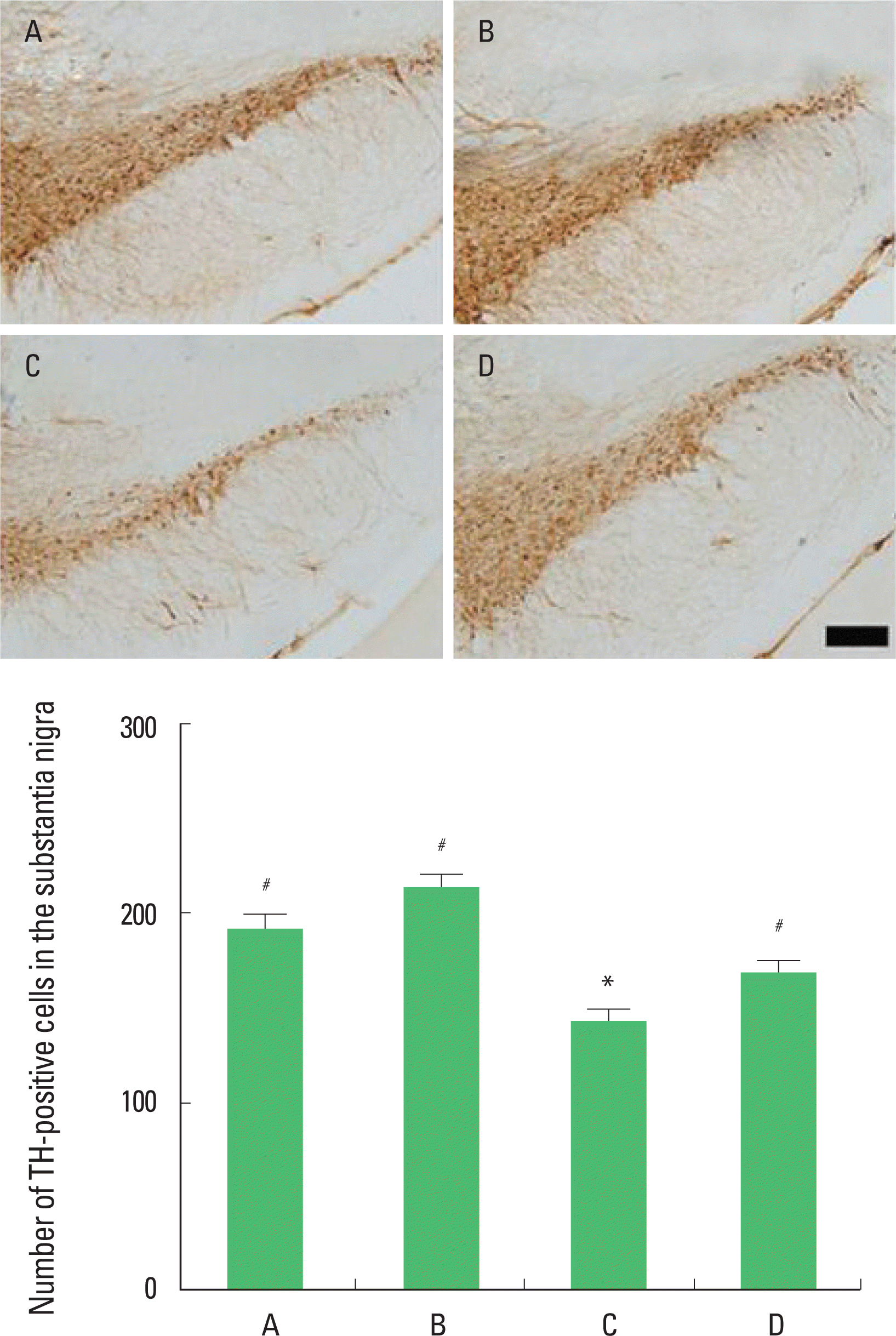
Effect of treadmill exercise on the TH expression in the substantia nigra
Photomicrographs of TH-positive cells in the substantia nigra are presented in Fig. 2. The number of TH-positive cells in the substantia nigra was 191.00±39.18/section in the in the sham-operation group, 213.73±31.24/section in the sham-operation and exercise group, 142.45±27.86/section in the hypoxic ischemia-induction group, and 168.70±23.97/section in the hypoxic ischemia-induction and exercise group. The present results show that induction of hypoxic ischemia brain injury decreased the number of TH-positive cells and that treadmill exercise increased the number of TH-positive cells in the hypoxic ischemia-induced rat pups.
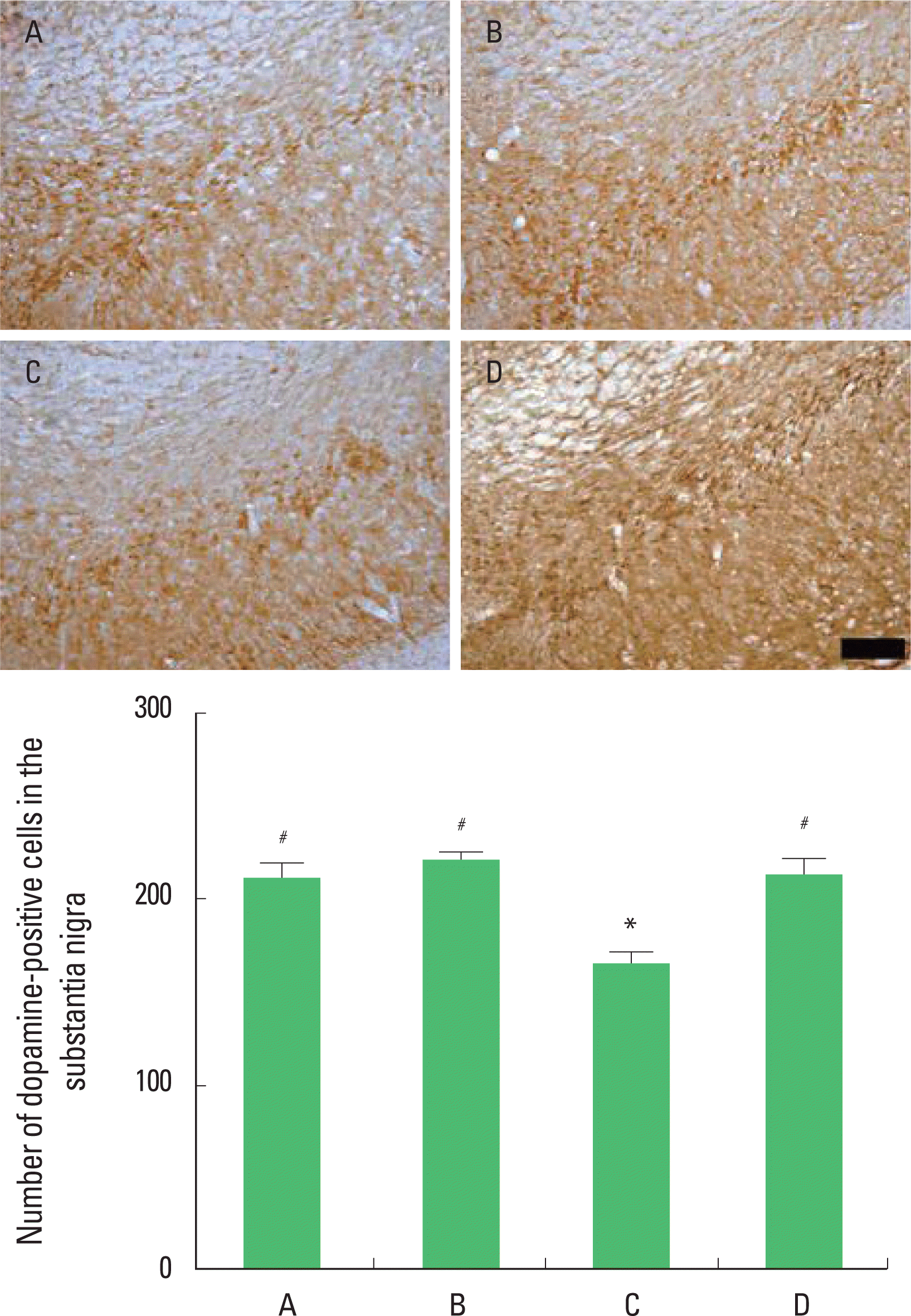
Effect of treadmill exercise on the dopamine expression in the substantia nigra
Photomicrographs of dopamine-positive cells in the substantia nigra are presented in Fig. 3. The number of dopamine-positive cells in the substantia nigra was 211.24±41.98/section in the in the sham-operation group, 220.41±25.37/section in the sham-operation and exercise group, 164.38±31.60/section in the hypoxic ischemia-induction group, and 210.96±43.14/section in the hypoxic ischemia-induction and exercise group. The present results show that induction of hypoxic ischemia brain injury decreased the number of dopamine-positive cells and that treadmill exercise increased the number of TH-positive cells in the substantia nigra in the hypoxic ischemia-induced rat pups.
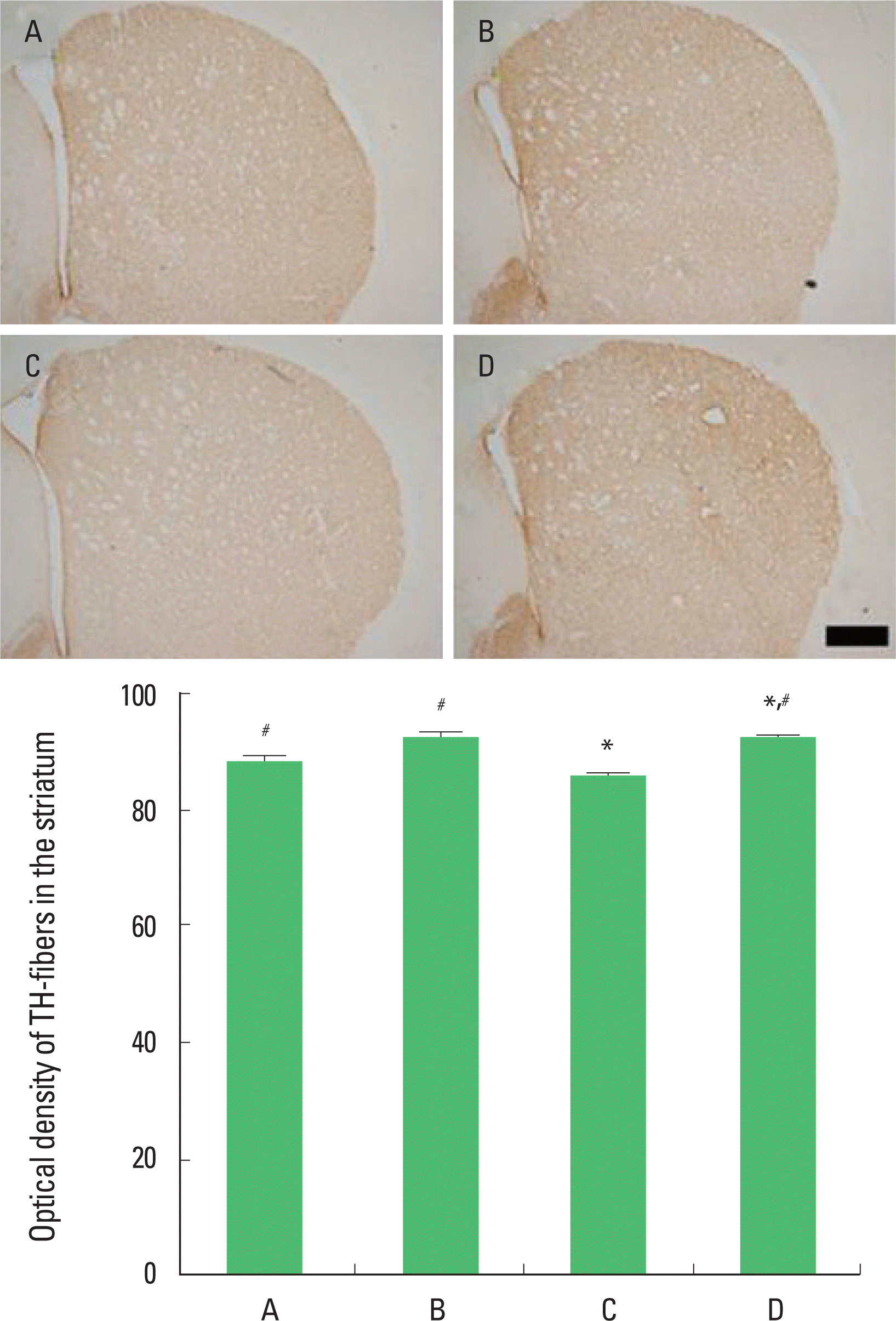
Effect of treadmill exercise on the TH expression in the striatum
Photomicrographs of TH-positive fibers in the striatum are presented in Fig. 4. The optical density of the TH-fibers in the striatum was 86.50±2.52/section in the in the sham-operation group, 90.46±3.83/section in the sham-operation and exercise group, 85.27±1.91/section in the hypoxic ischemia-induction group, and 89.29±3.59/section in the hypoxic ischemia-induction and exercise group. The present results that induction of hypoxic ischemia brain injury decreased the expression of TH-fibers and that treadmill exercise increased the expression of TH-fibers in the striatum in the hypoxic ischemia-induced rat pups.
DISCUSSION
Hypoxic ischemia brain injury occurring during the neonatal period leads to severe neurodevelopmental impairments such as cerebral palsy, which is associated with damage to the striatum (Brucknerová et al., 2006; Chalmers, 2005; Obenaus and Ashwal, 2008). Dopamine is implicated as an endogenous substance that mediates neuronal death after hypoxic ischemic insult (Adair and Filloux, 1992).
The present study showed that the hypoxic ischemia brain injury induced extensive degeneration of the dopaminergic neurons in the substantia nigra and a loss of dopaminergic fibers in the striatum. Hypoxic ischemia brain injury also deteriorated spatial learning ability in the Morris-water maze test. Burke et al. (1992) reported that neonatal hypoxic ischemia injury on postnatal 7 days resulted in a decrease in the TH-positive neurons in the substantia nigra of the neonatal and adult rats. The Rice-Vannucci model reduced the number of TH-positive cell bodies in the substantia nigra of the 4 weeks old rats (Oo et al., 1995). Fetal asphyxia caused profound loss of TH-positive neurons in the substantia nigra and TH-immunoreactivity in the striatum (Strackx et al., 2008). Hypoxic ischemia brain injury in the rodents also caused short-term and long-term learning deficits (Ikeda et al., 2001; Lubics et al., 2005).
In the present study, the reduction of TH-positive fibers in the striatum caused by the reduced number of TH-positive cells in the substantia nigra. The impairments in learning and memory following fetal hypoxic ischemia brain injury induced reduction in the number of TH-positive cell in the substantia nigra as well as a decreased TH-immunoreactivity in the striatum. Therefore, the altered TH activity in the present study may be part of the neural mechanisms contributing to the learning and memory impairment induced by neonatal hypoxic ischemia brain injury. These results might reflect impaired spatial learning ability via alteration of dopaminergic system.
In the present study, treadmill exercise ameliorated degeneration of the dopaminergic neurons in the substantia nigra and loss of dopaminergic fibers in the striatum. Following long-term tread-mill exercise, spatial learning ability was recovered near to the control level. Exercise stimulates dopamine synthesis in the striatum of epileptic mice (Sutoo and Akiyama, 2003). Exercise has been demonstrated to exert neuroprotective effects, enhance neurogenesis, and increase angiogenesis (Kim et al., 2013; Seo et al., 2013; Sung et al., 2012; Swain et al., 2003). Several neurotrophic factors might be involved in these beneficial effects of exercise (Yasuhara et al., 2007).
In the present results, treadmill exercise for 12 weeks alleviated hypoxic ischemia injury-induced disturbance of spatial learning ability through prevention of nigro-striatal dopaminergic neuronal loss. The present study suggests the possibility that treadmill exercise in early adolescent period may provide a useful strategy for the recovery after neonatal hypoxic ischemia brain injury.